- Equilibrio
electroquímico - Transporte activo
- Sistema de transporte activo
- Adenosintrifosfatasas del transporte
(ATPasas) - En la
superficie de F1 el ATP esta estabilizado frente al
ADP - Transporte activo de sodio y
potasio - Curvas
sigmoideas de flujo de iones en función de las
concentraciones - El
transporte activo de calcio
El flujo depende de la inclinación
de la curva de concentración. Donde la curva es más
inclinada se produce el mayor flujo.
Esta inclinación está
indicando cuanto cambia la concentración por unidad de
distancia.

Esta inclinación o pendiente de la
curva de concentración, se denomina "gradiente de
concentración".
La densidad de flujo es proporcional al
gradiente de concentración
Esta relación se denomina primera
Ley de Fick.
Dado un soluto S, a mayores gradientes de
concentración corresponden mayores densidades de flujo. Si
consideramos otro soluto T, molécula más grande.
Tendrá más dificultad para moverse, a iguales
gradientes corresponderán flujos menores de T que de S. A
ambos solutos le es aplicable la ley de Fick, pero se introduce
el concepto de difusibilidad.
Coeficiente de difusión
Unidad: Cm2/seg.
En el espesor de las membranas los
coeficientes de difusión son mucho menores que en el agua
para la mayoría de los solutos iónicos.
Si realizamos un gráfico de flujo en
función de la concentración, obtenemos:

Observamos una relación lineal entre
flujo y concentración, donde a través de su
pendiente podemos obtener la permeabilidad, la cual se define
como la capacidad de la membrana de permitir el paso de diversas
moléculas. Además, es una propiedad de la membrana
y del ion.
EQUILIBRIO
ELECTROQUÍMICO
El equilibrio electroquímico se
establece cuando las fuerzas eléctricas y las
químicas (debidas a diferencias de
concentración) son iguales y de sentido
contrario.

Para que un ion este en equilibrio
electroquímico debe haber una relación definida
entre las concentraciones y la diferencia de potencial
eléctrico entre ambos compartimentos.
TRANSPORTE
ACTIVO
El ion sodio se encuentra en una
situación particular. Por gradiente de
concentración y por atracción eléctrica
entra constantemente a la célula. Su potencial de
equilibrio es VIE = + 65 Mv.
Sin embargo, el valor del potencial de
reposo de la membrana es de – 90 mV.
El sodio entra a la célula
espontáneamente, a favor de un gradiente
electroquímico, por lo tanto se debe realizar trabajo para
extraerlo. Esto lo realiza la bomba sodio/ potasio, la cual
necesita la energía proveniente del metabolismo del ATP.
La misma requiere, potasio extracelular, sodio intracelular y ATP
intracelular.
A medida que aumenta la
concentración extracelular de potasio, aumenta la
velocidad de extracción de sodio por la
bomba.
El bombeo tiene una pequeña
corriente saliente, ya que según razones estequiometrias,
la bomba expulsa cada una molécula de ATP 3
Na+ e ingresa 2 K+.
Entonces, hacemos referencia a transporte
activo, aquel movimiento de soluto que se da gracias a una
reacción química, la cual permite que el soluto se
mueva en contra de su gradiente electroquímico.
SISTEMA DE
TRANSPORTE ACTIVO
El transporte activo se debe a la
existencia de sistemas especializados localizados en la membrana,
llamados sistemas de transporte activo o bombas de
cationes.
Propiedades:
· Los solutos sometidos a transporte
activo son cationes inorgánicos
pequeños.
· Los sistemas de transporte activo
están constituidos por proteínas asociados
íntimamente a la membrana.
· Los sistemas de transporte activo
poseen sitios a los que los cationes deben unirse para ser
transportados.
ADENOSINTRIFOSFATASAS DEL TRANSPORTE
(ATPasas)
Los sistemas de transporte activo usan como
fuente de energía la hidrólisis del
adenosíntrifosfato (ATP), adenosíndifosfato (ADP) y
fosfato inorgánico. Por este motivo, los sistemas poseen
poder catalítica de ATPasa, la que esta acoplada a la
transferencia de cationes, lo que conlleva a que se dé la
reacción, únicamente si hay presencia de los
cationes que la bomba transporta.
Las 2 clase de ATPasa del
transporte
ATPasa F0F1: Estas se ubican en la membrana
externa de células procariotas y en las membranas de la
mitocondrias, cloroplastos y vesículas secretorias de
vesículas eucariotas.
Estas acoplan reversiblemente la
transferencia de H+ a la hidrólisis de ATP.
Su nombre se debe a que posee 2 componentes: el
componente F1 se encuentra adosado a la superficie de la
membrana, pero no la penetra. Es la responsable de la actividad
ATPásica del sistema, pero por sí sola, es incapaz
de acoplar esta actividad con el transporte de H+.
El componente F0 está formado por
proteínas hidrofóbicas, que atraviesan la membrana
de lado a lado y que por uno de sus extremos se une a F1. No
posee actividad ATPasa, pero si permite la transferencia de
H+ a través de la
membrana.
Las membranas que poseen ATPasas FoF1
poseen también sistemas que catalizan reacciones de
oxidoreducción. Por ejemplo, la cadena respiratoria en la
membrana celular procariotas y en la membrana interna de las
mitocondrias. La energía liberada por las reacciones de
oxidoreducción se usa para impulsar un transporte activo
de H+, que da origen a una diferencia de potencial
electroquímico de H+ a ambos lados de la
membrana.
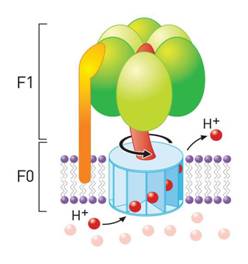
A modo de ejemplo, en la membrana
mitocondrial interna se encuentra una proteína formada por
estos dos tipos de constituyentes; la ATP sintasa.
La ATP sintasa mitocondrial cataliza la
formación de ATP a partir de ADP y fosfato
acompañada por el flujo de protones desde el lado P de la
membrana al lado N de la misma. El componente F1 es una
proteína periférica y el constituyente F0 es una
proteína integral (el subíndice significa que es
sensible a la oligomecina).
EN LA SUPERFICIE
DE F1 EL ATP ESTA ESTABILIZADO FRENTE AL ADP
En la superficie de la enzima, la
reacción ADP + Pi ? ATP + H20 es fácilmente
reversible: La energía libre de la síntesis de ATP
es cercana a cero. En solución, se necesita –
30
Kj/mol para hidrolizar el ATP.
¿A qué se debe esta gran
diferencia?
La ATP sintasa estabiliza el ATP con
respecto al DAP y al Pi uniéndolo más fuerte,
liberando la energía suficiente para contrarrestar el
efecto de fabricar ATP. La enzima, une ATP con mayor afinan que
ADP.
Es el gradiente protónico el que
provoca que la enzima libere el ATP de su
superficie.
ATPasas E1E2
Las más importantes son las
responsables del transporte activo de Na+
y K+ , de Ca+, de
H+ y K+. Durante su funcionamiento
adquieren conformaciones diferentes, las cuales difieren en
su aspecto funcional.
Las propiedades generales de estas ATPasas
transcurre en una serie de pasos elementales, que se pueden
enumerar de la siguiente forma:
1. El ATP se une al E1, que da lugar al E1
ATP. La unión se realiza en un sitio de unión en la
cara citoplasmática de la membrana.
2. El fosfato terminal del ATP se transfiere al
carboxilo beta de un residuo aspartilo de la enzima, con
formación de la fosfoenzima E1P con liberación de
ADP en el citoplasma. Para que ocurra esto, debe estar presente
el catión Mg2+. Este requerimiento es
común a todas las ATPasas E1E2, pero en ninguna de
ellas es transportado este catión.
3. Lo que ocurre es un cambio
conformacional de la enzima, la cual se transforma en
E2P.
Durante esta transición, el
catión es liberado al espacio extracelular.
4. La capacidad catalítica de E2P,
causa la hidrólisis de la fosfoenzima con la siguiente
liberación del fosfato inorgánico hacia el citosol
y formación de E2.
Cuando la ATPasa transporta dos cationes en
dirección opuesta, la defosforilación es acelerada
por el catión extracitoplasmático, el cual se
relaciona con la enzima en el espacio extracelular. Todas las
ATPasas son inhibidas por el orlovanadato (VO4), actuando sobre
el sitio donde se libera el fosforo.
5. Ocurre la reconversión de E2 en
E1. Este paso es el más lento, y por lo tanto, el que
limita la velocidad de la reacción global de
hidrólisis del ATP.
TRANSPORTE ACTIVO
DE SODIO Y POTASIO
Distribución de los iones entre el
citosol y el medio extracelular.
La concentración de
K+ en el citosol es de 150 mM, unas
30 veces mayor que la del medio extracelular, mientras que la de
Na+ es de 10 mM. En consecuencia,
existen gradientes de concentración que impulsan flujos
pasivos de Na+ hacia el interior y de
K+ hacia el exterior de la
célula.
La distribución de Na+ y K+ se
mantiene alejada del equlibrio debido a la acción de un
único sistema de transporte activo que induce flujos de
Na+ y K+ que balancean los flujos pasivos y mantiene así
la célula en estado estacionario ( estado de no
equilibrio, donde existe gasto de energía, ya que se va en
contra de un gradiente).
El transporte activo de Na+ y K+ esta
mediado por una ATPasa del tipo E1E2 llamada bomba de Na+,
K´ATPasa. Esta bomba está ubicada en la membrana
plasmática de todas las células eucariotas. En
células, donde el transporte es muy rápido, como
las de la rama gruesa del asa de Henle del
riñón, hasta el 25% del contenido de
proteínas en la membrana, por lo contrario en la membrana
de los eritrocitos se encuentra en una porcentaje de 0,1%. Esto
revela, que la velocidad de transporte activo de estos iones,
depende de la concentración en la membrana
plasmática de esta proteína integral.
La bomba sodio/potasio cataliza el
intercambio de 3 iones de sodio intracelular por 2 iones de
potasio extracelular, por cada molécula de ATP que es
hidrolizada.

Debido al número desigual de iones
que transporta la bomba, se genera una pequeña corriente
de cargas positivas hacia el exterior que incrementa la
electronegatividad en el interior celular. Por este motivo, se
dice que el intercambio es electrogénico
(Bombas iónicas que generan una diferencia
de potencial o carga eléctrica entre ambos lados de la
membrana celular, como las bombas de sodio-potasio).
CURVAS SIGMOIDEAS
DE FLUJO DE IONES EN FUNCIÓN DE LAS
CONCENTRACIONES
En condiciones fisiológicas, la
activación del intercambio de Na+
por K+, por Na+ intracelular
alcanza su valor semimáximo cuando la concentración
de sodio es de 10 a 15 mM. Esto lleva a que en condiciones
fisiológicas halla un 50% de unidades de bombas ocupadas
con el ion sodio, lo cual implica que pequeñas variaciones
en la concentración de sodio intracelular genere grandes
flujos de este ion, o en otras palabras genere una mayor
velocidad de transporte. El potasio intracelular actúa
como un inhibidor competitivo de la activación por sodio.
La bomba es altamente selectiva para el sodio intracelular, solo
el litio y el hidrógeno pueden sustituirlo.
La activación del intercambio de
sodio por potasio extracelular sigue una curva sigmoide, que para
concentraciones fisiológicas de sodio extracelular
(140-150 mM), alcanza su valor semimáximo cuando la
concentración de potasio es de 1 a 2 mM.
En condiciones fisiológicas, donde
la concentración de potasio es de 5 mM, un 70% de unidades
estarán combinadas con este catión.
El sodio extracelular actúa como un
inhibidor competitivo de la activación por
potasio.
Los sitios extracelulares son menos
selectivos que los intracelulares. El Rb+, litio, sodio y amonio
(NH4+) pueden ser transportados hacia el interior reemplazando el
potasio.
INHIBIDORES DE LA SODIO/POTASIO
ATPasa
Como todas las ATPasas E1E2, la bomba
sodio/potasio es inhibida por el vanadato. La enzima
también es inhibida por glucósidos
cardíacos, estos son inhibidores altamente
específicos de la bomba y ningún otro sistema de
transporte activo es sensible a ellos.
EL TRANSPORTE
ACTIVO DE CALCIO
El calcio citosólico actúa
como segundo mensajero que transmite la información del
estímulo a los sistemas que inician la respuesta. El
calcio puede cumplir con esta especial función, ya que su
concentración ciitosólica es muy baja, y flujos de
calcio generaran grandes cambios.
El calcio cumple numerosas funciones con
respecto a las respuestos celulares que media, como
son:
· Contracción
muscular
· Secreción de
glándulas endócrinas y exócrinas
· División celular
· Liberación de
neurotransmisores.
Una de las estrategias usada por las
células es la liberación de calcio estimulada por
calcio, donde un estímulo puede aumentar la
concentración de calcio citosólica facilitando el
ingreso de calcio extracelular, lo cual remueve calcio contenido
en organelos intracelulares, como lo son el retículo
sarcoplasmático del músculo.
El calcio penetra en la célula
gracias a componentes proteicos que se encuentran a nivel de la
membrana plasmática, son los canales de calcio
dependientes de voltaje, ya que en condiciones de reposo
permanecen cerrados, pero con la despolarización de la
membrana por un estímulo, estos censan ese cambio de
potencial y se abren permitiendo la entrada de calcio de forma
pasiva.
Los ejemplos más interesantes sobre
el papel del calcio, se radica a nivel del músculo, tanto
estriado como cardíaco, donde el calcio participa en el
mecanismo de la contracción muscular, función
fundamental para la supervivencia.
LAS Ca2+
ATPasas (retículo sarcoplasmático)
En condiciones fisiológicas, esta
bomba acopla el transporte de iones calcio a la hidrólisis
de ATP a ADP y Pi en la superficie citoplasmática de la
membrana. El transporte prosigue hasta que lo bloquea el efecto
inhibitorio del calcio que se acumula en el espacio
intrareticular. Entre el 70% y 80% de proteínas de
membrana del retículo corresponden a la bomba calcio
ATPasa.
Esto tiene vital importancia para tratar
con detalle la contracción muscular, ya que en pocas
palabras, luego de la contracción muscular se da la
relajación del músculo, y la misma se debe a la
rápida captación del calcio citosólico hacia
el retículo. Esto está dado, gracias a la bomba que
se encuentra en la membrana del retículo a un porcentaje
relativamente importante.
Calcio ATPasas de la membrana
celular
La membrana celular posee dos sistemas
capaces de inducir flujos activos de salida de calcio.
Son:
· Ca2+
ATPasa (presente en todas las células eucariotas, en
un 0,5%).
· Sistema de contratransporte que
acopla el flujo de salida de Ca2+ al
flujo de entrada de Na+.
En condiciones fisiológicas el
sistema acopla al transporte activo de calcio desde el citosol al
espacio extracelular a la hidrólisis de ATP a ADP y Pi en
la superficie citoplasmática de la membrana.
Autor:
Horacio Valdomir