Núcleo
E l núcleo celular es un orgánulo membranoso
que se encuentra en las células eucariotas. Contiene la
mayor parte del material genético celular, organizado en
múltiples moléculas lineales de ADN de gran
longitud formando complejos con una gran variedad de
proteínas como las histonas para formar los cromosomas. El
conjunto de genes de esos cromosomas se denomina genoma nuclear.
La función del núcleo es mantener la integridad de
esos genes y controlar las actividades celulares regulando la
expresión génica. Por ello se dice que el
núcleo es el centro de control de la célula.
Las principales estructuras que constituyen el núcleo
son la envoltura nuclear, una doble membrana que rodea
completamente al orgánulo y separa ese contenido del
citoplasma, además de contar con poros nucleares que
permiten el paso a través de la membrana para la
expresión genética y el mantenimiento
cromosómico.
- a. Envoltura nuclear: La envuelta nuclear
separa físicamente al nucleoplasma (cromatina y
demás componentes del interior nuclear) del citoplasma
y regula el movimiento de macromoléculas entre ellos,
establece la forma nuclear y contribuye a la
organización interna del núcleo ya que aporta
lugares de anclaje para la cromatina. La envuelta nuclear
está formada por una membrana doble, externa e
interna, quedando entre ambas un espacio intermembranoso de
aproximadamente 25-40 nm, formando todos estos elementos las
denominas cisternas perinucleares. La membrana externa se
continúa con la del retículo
endoplasmático y posee ribosomas adheridos. La
membrana interna contiene una composición molecular
diferente y posee proteínas transmembrana que
interactúan con la cromatina y con la lámina
nuclear, el otro componente de la envuelta nuclear. Existe
una comunicación entre la membrana nuclear interna y
externa en la periferia de los poros nucleares. Las
proteínas se sintetizan en el retículo
endoplasmático que llegan a la membrana interna por
difusión, pero sólo aquellas que interaccionan
con las proteínas de la lámina nuclear o de la
cromatina se mantienen aquí. - b. Cromatina: El nucleoplasma, rodeado por la
envuelta nuclear, contiene la cromatina, la cual se puede
considerar como el ADN (ácido desoxirribonucleico)
más todas las moléculas relacionadas con su
organización, fundamentalmente histonas. El ADN
está formado por 4 desoxirribonucleótidos
(abreviado como nucleótidos). Cada nucleótido
contiene una sucesión de tres componentes: base,
pentosa y grupo fosfato. Las bases son cuatro, dos
púricas: adenina (A) y guanina (G), y dos
pirimidínicas: timina (T) y citosina (C). La pentosa
es la desoxiribosa. Cada base se une a una pentosa formando
un desoxinucleósido. Cada desoxirribonucleósido
se une un grupo fostato por un carbono de la pentosa
formándose un desoxirribonucleótido.
Así, una cadena de ADN está formado por una
sucesión de nucleótidos unidos entre sí
por los grupos fosfato. Esto es una cadena simple pero el ADN
está formado por dos cadenas simples gracias a la
complementariedad que existe entre las bases A y T y entre G
y C, las cuales establecen uniones del tipo puentes de
hidrógeno. Las dos hebras son antiparalelas, es decir,
que en los extremos tenemos el carbono 3' de una cadena y el
5' de la otra. Ambas se disponen en forma de doble
hélice de unos 2.5 nm de anchura. Los
nucleótidos no sólo están en el ADN.
Pueden estar formando parte de otras moléculas con
funciones totalmente diferentes. Por ejemplo el ATP
(adenosín trifosfato) es la molécula de
transferencia energética, o el AMPc (adenosín
monofostato cíclico) que es un segundo mensajero
celular muy importante. El ADN no se encuentra libre en el
núcleo sino asociado a proteínas como las
histonas y a otras proteínas implicadas en su
procesamiento, formando en conjunto la cromatina. Las
histonas son proteínas asociadas al ADN que determinan
su organización. . Es interesante señalar que
cuando se ha producido la división celular y se
vuelven a desempaquetar a los cromosomas, la cromatina de
cada uno de ellos suele ocupar un territorio concreto dentro
del interior nuclear. Es decir, en el interior del
núcleo no existe una madeja enredada de cromatina
correspondiente a cromosomas diferentes sino un espacio
compartimentado donde cada cromosoma suele ocupar una
región del nucleoplasma más o menos delimitada
donde cada cromosoma descondensa su cromatina.

- c. Nucléolo: El nucléolo es un
compartimento nuclear formado por cromatina y visible al
microscopio óptico. Las células de
mamíferos contienen desde 1 a 5 nucléolos. Sus
dimensiones varían dependiendo de la actividad de la
célula y puede llegar a ser muy grande, del orden de
micrómetros de diámetro. Normalmente las
células que están realizando una gran
síntesis proteica poseen nucléolos grandes.
Durante la mitosis desaparece, permitiendo a la cromatina que
lo forma reorganizarse para constituir los cromosomas. En el
nucléolo se dan procesos relacionados con la
generación de los ribosomas: síntesis y
maduración del ARN ribosómico (ARNr) y
ensamblaje de las subunidades ribosómicas. El
ensamblaje de las subunidades ribosómicas es un
proceso curioso de trasiego de moléculas entre el
citoplasma y el nucleoplasma. Primero se transcriben los
genes de dichas proteínas, que se localizan fuera de
la cromatina nucleolar. Éste ARNm debe salir al
citosol donde es traducido a proteínas por los
ribosomas libres. Estas proteínas entrarán en
el núcleo y llegan hasta el nucléolo.
Aquí se asocian con los ARNr para formar las
subunidades ribosómicas que deberán ser
exportadas de nuevo al citosol atravesando otra vez los poros
nucleares. Así, la visibilidad del nucléolo se
debe a que muchos genes que producen ARNr se están
transcribiendo, a que hay muchas proteínas implicadas
en el procesamiento de ese primer transcrito, a las
proteínas de las subunidades ribosómicas y a
aquellas proteínas relacionadas con el ensamblaje de
éstos. Se estima que hay unas 690 proteínas
diferentes asociadas de forma estable con el
nucléolo.

- d. Nucleoesqueleto: Las lamininas constituyen
los principales componentes de nucleoesqueleto, son las
determinantes del tamaño nuclear, del mantenimiento de
la forma nuclear, de la integridad mecánica del
núcleo, proporcionando apoyo estructural a la
periferia nuclear, así como del espacio existente
entre los distintos complejos nuclear del poro. Las lamininas
actúan también en la organización de la
cromatina al actuar como sitios de anclaje o de unión
para la cromatina. Una matriz de láminas se extiende
hacia el interior del núcleo. La cromatina contenida
en un núcleo de una célula en interface se
organiza en grandes bucles de DNA y regiones
específicas de estos bucles se unen a la matriz de
láminas. La organización laminar normal es
esencial para la replicación del DNA y puede jugar un
papel en la regulación de la expresión
génica. Las láminas tienen también un
papel fundamental durante reorganización del
núcleo en la división celular. Así, la
lámina nuclear participa en el control del
desensamblaje del núcleo en la profase y la
reorganización de nuevo del mismo en la telofase
durante el proceso de mitosis
Orgánulos
- a. Ribosomas: Los ribosomas, visibles
al microscopio electrónico como
partículas esféricas, son complejos
supramoleculares encargados de
ensamblar proteínas a partir de la
información genética que les llega del ADN
transcrita en forma de ARN mensajero. Elaborados en
el núcleo, desempeñan su función de
síntesis de proteínas en el citoplasma.
Están formados por ARN ribosómico y
por diversos tipos de proteínas. Estructuralmente,
tienen dos subunidades. En las células, estos
orgánulos aparecen en diferentes estados
de disociación. Cuando están completos,
pueden estar aislados o formando grupos (polisomas).
También pueden aparecer asociados
al retículo endoplasmático rugoso o a
la envoltura nuclear. Los ribosomas son estructuras
globulares, carentes de membrana. Pueden encontrarse libres
en el citoplasma o adheridos a las membranas del
retículo endoplasmático. Unas proteínas
(riboforinas) sirven de nexo entre ambas estructuras. Su
estructura es sencilla: dos subunidades (una mayor o otra
menor) de diferente coeficiente de sedimentación. Su
función consiste únicamente en ser el
orgánulo lector del ARN mensajero, con órdenes
de ensamblar los aminoácidos que formarán la
proteína. Son orgánulos sintetizadores de
proteínas.
- b. Retículo endoplasmático: El
retículo endoplasmático es orgánulo
vesicular interconectado que forma cisternas, tubos aplanados
y sáculos comunicados entre sí. Intervienen en
funciones relacionadas con la síntesis proteica,
glicosilación de
proteínas, metabolismo de lípidos y
algunos esteroides, detoxificación,
así como el tráfico de vesículas.
En células especializadas, como
las miofibrillas o células musculares, se
diferencia en el retículo sarcoplásmico,
orgánulo decisivo para que se produzca
la contracción muscular. El retículo
endoplasmático rugoso se encuentra unido a la membrana
nuclear externa mientras que el retículo
endoplasmático liso es una prolongación del
retículo endoplasmático rugoso. El
retículo endoplasmático rugoso tiene esa
apariencia debido a los numerosos ribosomas adheridos a su
membrana mediante unas proteínas denominadas
"riboforinas". Tiene unos sáculos más
redondeados cuyo interior se conoce como "luz del
retículo" o "lumen" donde caen las proteínas
sintetizadas en él. Está muy desarrollado en
las células que por su función deben realizar
una activa labor de síntesis, como las células
hepáticas o las células del páncreas. El
retículo endoplasmático liso no tiene ribosomas
y participa en el metabolismo de lípidos. El
retículo endoplasmático tiene variedad de
formas: túbulos, vesículas, cisternas. En
algunos casos en una misma célula se pueden observar
los tres tipos.

- c. Aparato de Golgi: es un orgánulo
formado por apilamientos de sáculos
denominados dictiosomas, si bien, como ente
dinámico, estos pueden interpretarse como estructuras
puntuales fruto de la coalescencia de
vesículas. Recibe las vesículas
del retículo endoplasmático
rugoso que han de seguir siendo procesadas. Dentro de
las funciones que posee el aparato de Golgi se encuentran
la glicosilación de proteínas,
selección, destinación, glicosilación
de lípidos y la síntesis
de polisacáridos de la matriz extracelular.
Posee tres compartimientos; uno proximal al retículo
endoplasmático, denominado
«compartimento cis», donde se
produce la fosforilación de las manosas de
las enzimas que han de dirigirse al lisosoma; el
«compartimento intermedio», con
abundantes manosidasas y N-acetil-glucosamina transferasas;
y el «compartimento o red trans»,
el más distal, donde se transfieren residuos
de galactosa y ácido siálico, y
del que emergen las vesículas con los diversos
destinos celulares.
Las principales funciones del aparato de Golgi vienen a ser
las siguientes:
- Modificación de sustancias sintetizadas en el RER:
En el aparato de Golgi se transforman las sustancias
procedentes del RER. Estas transformaciones pueden ser
agregaciones de restos de carbohidratos para conseguir la
estructura definitiva o para ser proteolizados y así
adquirir su conformación activa. Por ejemplo, en el
RER de las células acinosas del páncreas se
sintetiza la proinsulina que debido a las transformaciones
que sufre en el aparato de Golgi, adquirirá la forma o
conformación definitiva de la insulina. Las enzimas
que se encuentran en el interior de los dictiosomas son
capaces de modificar las macromoléculas mediante
glicosilación (adición de carbohidratos) y
fosforilación (adición de fosfatos). Para ello,
el aparato de Golgi transporta ciertas sustancias como
nucleótidos y azúcares al interior del
orgánulo desde el citoplasma - Secreción celular: las sustancias atraviesan todos
los sáculos del aparato de Golgi y cuando llegan a la
cara trans del dictiosoma, en forma de vesículas de
secreción, son transportadas a su destino fuera de la
célula, atravesando la membrana citoplasmática
por exocitosis. Un ejemplo de esto son los proteoglicanos que
conforman la matriz extracelular de los animales. El aparato
de Golgi es el orgánulo de mayor síntesis de
carbohidratos.5 Esto incluye la producción de
glicosaminoglicanos (GAGs), largos polisacáridos que
son anclados a las proteínas sintetizadas en el RE
para dar lugar a los proteoglicanos. De esto se
encargarán las enzimas del Golgi por medio de un
residuo de xilosa. - Producción de membrana plasmática: los
gránulos de secreción cuando se unen a la
membrana en la exocitosis pasan a formar parte de esta,
aumentando el volumen y la superficie de la
célula. - Formación de los lisosomas primarios.
- Formación del acrosoma de los espermios.

- d. Lisosomas:
son orgánulos que albergan multitud de
enzimas hidrolíticas. De morfología muy
variable, no se ha demostrado su existencia en células
vegetales. Una característica que agrupa a todos
los lisosomas es la posesión de hidrolasas
ácidas: proteasas, nucleasas,
glucosidasas, lisozima, arilsulfatasas,
lipasas, fosfolipasas y fosfatasas.
Procede de la fusión de vesículas procedentes
del aparato de Golgi, que, a su vez, se fusionan en un tipo
de orgánulo denominado endosoma temprano, el
cual, al acidificarse y ganar en enzimas hidrolíticos,
pasa a convertirse en el lisosoma funcional. Sus funciones
abarcan desde la degradación de macromoléculas
endógenas o procedentes de la fagocitosis a
la intervención en procesos de apoptosis. El pH
en el interior de los lisosomas es de 4,8 (bastante menor que
el del citosol, que es neutro) debido a que las enzimas
proteolíticas funcionan mejor con un pH ácido.
La membrana del lisosoma estabiliza el pH bajo bombeando
iones (H+) desde el citosol, y así mismo, protege al
citosol e igualmente al resto de la célula de las
enzimas digestivas que hay en el interior del lisosoma. Las
enzimas lisosomales son capaces de digerir bacterias y otras
sustancias que entran en la célula por fagocitosis, u
otros procesos de endocitosis. Los lisosomas utilizan sus
enzimas para reciclar los diferentes orgánulos de la
célula, englobándolos, digiriéndolos y
liberando sus residuos en el citosol. De esta forma los
orgánulos de la célula se están
continuamente reponiendo. El proceso de digestión de
los orgánulos se llama autofagia. Por ejemplo, las
células hepáticas se reconstituyen por completo
una vez cada dos semanas. Las enzimas más importantes
del lisosoma son: Lipasas, que digiere lípidos;
Glucosidasas, que digiere carbohidratos; Proteasas, que
digiere proteínas; Nucleasas, que digiere
ácidos nucleícos.

e. Cuerpos multivesiculares: Los cuerpos
multivesiculares y posteriormente los endosomas
tardíos son la antesala de la degradación de
las moléculas endocitadas, la cual se realiza
finalmente en los lisosomas gracias a unas enzimas
denominadas hidrolasas ácidas. Las moléculas
destinadas a la degradación llegan desde los endosomas
tempranos (bien mediante vesículas o bien mediante la
transformación de los endosomas tempranos en cuerpos
multivesiculares). Las hidrolasas ácidas
también llegan a los endosomas tardíos
empaquetadas en vesículas enviadas desde el TGN del
aparato de Golgi. Desde éstos se producirá un
último reciclado mediante vesículas hacia
endosomas tempranos y hacia el TGN del aparato de Golgi. Sin
embargo, estas enzimas no tendrán su máxima
actividad hasta llegar a los lisosomas. Desde los endosomas
tardíos se produce un último reciclado de
vesículas hacia el TGN y endosomas tempranos. La
acción de las bombas de protones localizadas en las
membranas de estos endosomas irá acidificando
progresivametne el pH interno y por tanto favoreciendo la
acción de las hidrolasas ácidas, cuya actividad
óptima se da a un pH próximo a 5, el cual se
alcanza en los lisosomas. El aspecto multivesicular que se
observa a microscopía electrónica de los
cuerpos multivesiculares se debe a que en sus membranas se
producen invaginaciones que resultarán en
vesículas en su interior. De esta manera se pueden
degradar las moléculas que forman parte integral de
las membranas, aunque en dichas invaginaciones entra
además parte del fluido citosólico, que
también será degradado. Como dijimos
anteriormente los endosomas tardíos se forman por
maduración de los cuerpos multivesiculares. Algunos
tipos celulares como las células
hematopoyéticas, los linfocitos, las células
dendríticas, las células epiteliales
intestinales, los mastocitos y las células tumorales,
realizan un tipo de tráfico vesicular un tanto
extraño. Los cuerpos multivesiculares, en vez de
convertirse en lisosomas, se fusionan con la membrana
plasmática liberando sus vesículas internas (de
30 a 60 nm de diámetro) al espacio extracelular. A
estas vesículas liberadas se les denomina exosomas y
poseen una composición molecular distinta a otros
compartimentos intracelulares, por ejemplo poseen mucho
colesterol y esfingomielina.

f. Laminillas anulares: son
orgánulos poco frecuentes que aparecen, por ejemplo,
en las células que se dividen con rapidez, como las
células germinales en sus primeras etapas y algunas
células cancerosas. Consisten en pilas o rimeros de
cisternas limitadas por membrana, de forma aplanada y
disposición paralela, en cuya superficie aparecen
poros. Esto recuerda la envoltura nuclear con sus poros y se
cree que estas membranas anulares son precursoras de
envolturas nucleares o sitios de almacenamiento de
RNA.

g. Peroxisomas: Los peroxisomas son
orgánulos muy comunes en forma de vesículas que
contienen abundantes enzimas de tipo oxidasa y catalasa; de
tan abundantes, es común que cristalicen en su
interior. Estas enzimas cumplen funciones de
detoxificación celular. Otras funciones de los
peroxisomas son: las oxidaciones flavínicas generales,
el catabolismo de las purinas, la beta-oxidación de
los ácidos grasos, el ciclo del glioxilato, el
metabolismo del ácido glicólico y la
detoxificación en general. Se forman de
vesículas procedentes del retículo
endoplasmático.

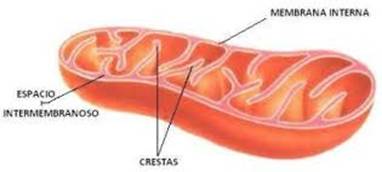
h. Mitocondrias: Las mitocondrias son
orgánulos de aspecto, número y tamaño
variable que intervienen en el ciclo de
Krebs, fosforilación oxidativa y en
la cadena de transporte de electrones de
la respiración. Presentan una doble membrana,
externa e interna, que dejan entre ellas un espacio
perimitocondrial; la membrana interna, plegada en crestas
hacia el interior de la matriz mitocondrial, posee una
gran superficie. En su interior posee generalmente una sola
molécula de ADN, el genoma mitocondrial,
típicamente circular, así como ribosomas
más semejantes a los bacterianos que a los
eucariotas.12 Según la teoría
endosimbiótica, se asume que la primera
protomitocondria era un tipo de proteobacteria. La
principal función de las mitocondrias es la
oxidación de metabolitos (ciclo de Krebs,
beta-oxidación de ácidos grasos) y la
obtención de ATP mediante la fosforilación
oxidativa, que es dependiente de la cadena transportadora de
electrones; el ATP producido en la mitocondria supone un
porcentaje muy alto del ATP sintetizado por la célula.
También sirve de almacén de sustancias como
iones, agua y algunas partículas como restos de virus
y proteínas. La morfología de la mitocondria es
difícil de describir puesto que son estructuras muy
plásticas que se deforman, se dividen y fusionan.
Normalmente se las representa en forma alargada. Su
tamaño oscila entre 0,5 y 1 µm de
diámetro y hasta 7 µm de longitud. Su
número depende de las necesidades energéticas
de la célula. Al conjunto de las mitocondrias de la
célula se le denomina condrioma celular.

i. Melanosomas: es un orgánulo
que contiene melanina, el pigmento absorbente de luz
más común en el reino animal. Las
células que producen melanosomas se denominan
melanocitos, mientras que las células que simplemente
han ingerido los melanosomas se denominan melanofagos. Los
melanosomas están delimitados por una membrana
lipídida y son generalmente esféricos o
alargados. Su forma es constante para un tipo dado de especie
y célula. Tienen una ultraestructura
característica en la microscopia electrónica,
que varía según la madurez del melanosoma.
Antes de que contenga los suficientes pigmentos para ser
visto por el microscopio óptico se conoce como
pre-melanosoma. En algunos melanocitos, los melanosomas
permanecen estáticos dentro de la célula. En
otros tipos de melanocitos, la célula puede extender
su superficie con seudópodos largos, llevando los
melanosomas lejos del centro de la célula y aumentando
la eficacia de la célula en la absorción de luz
absorbente. Por ejemplo, esto sucede lentamente en los
melanocitos cutáneos en respuesta a la luz
ultravioleta, a la vez que la producción de nuevos
melanosomas y de la donación creciente de melanosomas
a los queratinocitos adyacentes, las células normales
de la superficie de la piel. Estos cambios son colectivamente
responsables del bronceado después de la
exposición a la luz del sol o a los rayos
ultravioletas. En muchas especies de peces, anfibios,
crustáceos y reptiles, los melanosomas pueden ser
altamente móviles dentro de la célula en
respuesta al control hormonal (o a veces de los nervios), y
esto conduce a los cambios visibles de color que utilizan
para señalar su comportamiento. Los bonitos y
rápidos cambios de color de muchos cefalópodos
(pulpos y calamares) se basan sin embargo en un sistema
distinto, los cromatóforo. La melanina es una familia
de grandes polímeros sintetizados por un sistema de
enzimas, especialmente la tirosinasa). Se piensa que la
polimerización de la melanina tiene lugar por
amiloidogénesis de la proteína pMel, que
está presente en grandes cantidades en los
melanosomas.

Inclusiones
Consiste en sustancias nutritivas almacenadas, productos
secundarios inactivos del metabolismo o acumulaciones de
sustancias endógenas o exógenas que exhiben una
tinción (pigmentación) propia.
a. Partículas de
glucógeno: Forman parte de las inclusiones por
almacenamiento. Son depósitos de glucosa, que se
observan como partículas o rosetas fuertemente
electrón denso. A nivel de MO son PAS positivas. En
algunas células forman grandes agregados de
partículas.

b. Gotitas de lípidos: Los
lípidos biológicos constituyen un grupo
químicamente diversos de compuestos, cuya
característica común y definitoria es su
insolubilidad en agua. Las funciones biológicas de los
lípidos son igualmente diversas. En muchos organismos
las grasas y los aceites son las formas principales de
almacenamiento energético, mientras que los
fosfolípidos y los esteroles constituyen la mitad de
la masa de las membranas biológicas. Otros
lípidos, aun estando presentes en cantidades
relativamente pequeñas, juegan papeles cruciales como
cofactores enzimáticos, transportadores
electrónicos, agentes emulsionantes, hormonas y
mensajeros intracelulares.

c. Inclusiones cristalinas: En el
citoplasma existen sustancias inertes hidrofobicas llamadas
inclusiones. Se encuentran en todas las células
eucariotas ,tanto vegetales como animales, siendo las
inclusiones más comunes las de almidon y glucogeno.
Inclusiones cristalinas: se observan en forma de cristales, y
se trata de depósitos proteicos.
1. celulas vegetales: estas inclusiones proceden de
sales cristalizadas, formando cristales llamados drusas y
ráfides.
2. celulas animales: las células de los tubos
seminíferos de los mamíferos presentan inclusiones
llamadas cristales de Charcot Bottcher. Inclusiones
hidófobas: se trata de productos sintetizados por la
célula. 1. celulas vegetales: se encuentran en grandes
vacuolas o por el citoplasma. ejem: granos de almidón,
gotas de grasa, aceites esenciales, látex.. 2. celulas
animales: glucógeno: muy abundante en células
hepáticas y musculares.las células animales
utilizan el glucógeno que se ha acumulado en el
hígado como principal fuente energética.
Citoesqueleto..
Las células poseen un andamiaje que permite el
mantenimiento de su forma y estructura, pero más
aún, este es un sistema dinámico que
interactúa con el resto de componentes celulares generando
un alto grado de orden interno. Dicho andamiaje está
formado por una serie de proteínas que se agrupan dando
lugar a estructuras filamentosas que, mediante otras
proteínas, interactúan entre ellas dando lugar a
una especie de retículo. El mencionado andamiaje recibe el
nombre decitoesqueleto, y sus elementos mayoritarios son: los
microtúbulos, los microfilamentos y los filamentos
intermedios.
a. Microfilamentos: Los microfilamentos
o filamentos de actina están formados por
una proteína globular, la actina, que puede
polimerizar dando lugar a estructuras filiformes. Dicha
actina se expresa en todas las células del cuerpo y
especialmente en las musculares ya que está
implicada en la contracción muscular, por
interacción con la miosina. Además, posee
lugares de unión a ATP, lo que dota a sus
filamentos de polaridad. Puede encontrarse en forma
libre o polimerizarse en microfilamentos, que son
esenciales para funciones celulares tan importantes como la
movilidad y la contracción de la célula durante
la división celular.

Citoesqueleto eucariota: microfilamentos
en rojo, microtúbulos en verde y núcleo en
azul.
b. Microtúbulos: Los
microtúbulos son estructuras tubulares de 25 nm
de diámetro exterior y unos 12 nm de
diámetro interior, con longitudes que varían
entre unos
pocos nanómetros a micrómetros,
que se originan en los centros organizadores de
microtúbulos y que se extienden a lo largo de
todo el citoplasma. Se hallan en las células
eucariotas y están formadas por la
polimerización de un dímero de
dos proteínas globulares,
la alfa y la beta tubulina. Las tubulinas
poseen capacidad de unir GTP. Los
microtúbulos intervienen en diversos procesos
celulares que involucran desplazamiento
de vesículas de secreción,
movimiento de orgánulos, transporte intracelular
de sustancias, así como en la división celular
(mitosis y meiosis) y que, junto con
los microfilamentos y los filamentos
intermedios, forman el citoesqueleto. Además,
constituyen la estructura interna de los cilios y
los flagelos.c. Filamentos intermedios: Los
filamentos intermedios son componentes del citoesqueleto.
Formados por agrupaciones de proteínas fibrosas, su
nombre deriva de su diámetro, de 10 nm, menor que
el de los microtúbulos, de 24 nm, pero mayor
que el de los microfilamentos, de 7 nm. Son ubicuos
en las células animales, y no existen
en plantas ni hongos. Forman un grupo
heterogéneo, clasificado en cinco familias:
las queratinas, en células epiteliales;
los neurofilamentos, en neuronas;
los gliofilamentos, en células gliales;
la desmina, en músculo
liso y estriado; y la vimentina, en
células derivadas
del mesénquima.

Micrografía al microscopio
electrónico de barrido mostrando la superficie de
células ciliadas del epitelio de
los bronquiolos.
d. Centríolos: son una pareja de
estructuras que forman parte del citoesqueleto de
células animales. Semejantes a cilindros huecos,
están rodeados de un material proteico denso
llamado material pericentriolar; todos ellos forman
el centrosoma o centro organizador de
microtúbulos que permiten la
polimerización de microtúbulos de
dímeros de tubulina que forman parte del
citoesqueleto. Los centríolos se posicionan
perpendicularmente entre sí. Sus funciones son
participar en la mitosis, durante la cual generan
el huso acromático, y en
la citocinesis, así como, se postula,
intervenir en la nucleación de
microtúbulos.e. Cilios y flagelos: Se
trata de especializaciones de la superficie celular con
motilidad; con una estructura basada en agrupaciones de
microtúbulos, ambos se diferencian en la mayor
longitud y menor número de los flagelos, y en la mayor
variabilidad de la estructura molecular de estos
últimos.
Ciclo
celular.
a. Fases del ciclo celular.
El ciclo celular es el proceso ordenado y repetitivo en
el tiempo mediante el cual una célula madre
crece y se divide en dos células hijas. Las
células que no se están dividiendo se encuentran en
una fase conocida como G0, paralela al ciclo. La
regulación del ciclo celular es esencial para el correcto
funcionamiento de las células sanas, está
claramente estructurado en fases
El estado de no división o interfase. La
célula realiza sus funciones específicas y, si
está destinada a avanzar a la división celular,
comienza por realizar la duplicación de
su ADN.El estado de división, llamado fase M,
situación que comprende
la mitosis y citocinesis. En algunas
células la citocinesis no se produce,
obteniéndose como resultado de la división una
masa celular plurinucleada
denominada plasmodio.
A diferencia de lo que sucede en la mitosis, donde
la dotación genética se mantiene, existe una
variante de la división celular, propia de las
células de la línea germinal,
denominada meiosis. En ella, se reduce la dotación
genética diploide, común a todas
las células somáticas del organismo, a
una haploide, esto es, con una sola copia del genoma.
De este modo, la fusión, durante
la fecundación, de dos gametos haploides procedentes
de dos parentales distintos da como resultado un zigoto, un
nuevo individuo, diploide, equivalente en dotación
genética a sus padres.
La interfase consta de tres estadios claramente
definidos.
Fase G1: es la primera fase del ciclo celular, en la
que existe crecimiento celular con síntesis
de proteínas y de ARN. Es el
período que trascurre entre el fin de una mitosis y el
inicio de la síntesis de ADN. En él la
célula dobla su tamaño y masa debido a la
continua síntesis de todos sus componentes, como
resultado de la expresión de los genes que
codifican las proteínas responsables de
su fenotipo particular.Fase S: es la segunda fase del ciclo, en la que se
produce la replicación o síntesis del ADN.
Como resultado cada cromosoma se duplica y queda
formado por dos cromátidas idénticas.
Con la duplicación del ADN,
el núcleo contiene el doble de
proteínas nucleares y de ADN que al
principio.Fase G2: es la segunda fase de crecimiento del ciclo
celular en la que continúa la síntesis de
proteínas y ARN. Al final de este período se
observa al microscopio cambios en la estructura celular, que
indican el principio de la división celular. Termina
cuando los cromosomas empiezan a condensarse al inicio de la
mitosis.
La fase M es la fase de la división celular en la
cual una célula progenitora se divide en dos
células hijas hijas idénticas entre sí y a
la madre. Esta fase incluye la mitosis, a su vez dividida
en: profase, metafase, anafase, telofase; y
la citocinesis, que se inicia ya en la telofase
mitótica.
La incorrecta regulación del ciclo celular puede
conducir a la aparición de células
precancerígenas que, si no son inducidas al suicidio
mediante apoptosis, puede dar lugar a la aparición
de cáncer. Los fallos conducentes a dicha
desregulación están relacionados con
la genética celular: lo más común
son las alteraciones en oncogenes, genes supresores de
tumores y genes de reparación del
ADN.

b. Células madres y
células hijas
Existen diferentes tipos de células madre, aunque
las más empleadas en biología son las
células madre embrionarias y las adultas:
Células madre embrionarias (pluripotentes):
Generalmente se obtienen de la masa celular interna del
blastocisto. El blastocisto está formado por una capa
externa denominada trofoblasto, formada por unas 70
células, y una masa celular interna constituida por
unas 30 células que son las células madre
embrionarias que tienen la capacidad de diferenciarse en
todos los tipos celulares que aparecen en el organismo
adulto, dando lugar a los tejidos y órganos. En la
actualidad se utilizan como modelo para estudiar el
desarrollo embrionario y para entender cuáles son los
mecanismos y las señales que permiten a una
célula pluripotente llegar a formar cualquier
célula plenamente diferenciada del organismo.
Asimismo, están comenzando a ser utilizadas con
éxito en terapias biomédicas.Células madre germinales: Se trata de
células madre embrionarias pluripotenciales que se
derivan de los esbozos gonadales del embrión. Estos
esbozos gonadales se encuentran en una zona específica
del embrión denominada cresta gonadal, que dará
lugar a los óvulos y espermatozoides. Tienen una
capacidad de diferenciación similar a las de las
células madre embrionarias, pero su aislamiento
resulta más difícil.Células madre fetales: Estas células
madre aparecen en órganos fetales como,hígado,
pulmón y poseen características similares a sus
homólogas en tejidos adultos, aunque parecen mostrar
mayor capacidad de expansión y diferenciación.
Su procedencia no está del todo clara. Podrían
tener origen embrionario o bien tratarse de nuevas oleadas de
progenitores sin relación con las células madre
embrionarias.Células madre adultas: Son células no
diferenciadas que se encuentran en tejidos y órganos
adultos y que poseen la capacidad de diferenciarse para dar
lugar a células adultas del tejido en el que se
encuentran, por lo tanto se consideran células
multipotenciales. En un individuo adulto se conocen hasta
ahora alrededor de 20 tipos distintos de células
madre, que son las encargadas de regenerar tejidos en
continuo desgaste (como la piel o la sangre) o dañados
(como el hígado). Su capacidad es más limitada
para generar células especializadas. Las
células madre hematopoyéticas de médula
ósea (encargadas de la formación de la sangre)
son las más conocidas y empleadas en la clínica
desde hace tiempo. En la misma médula, aunque
también en sangre del cordón umbilical, en
sangre periférica y en la grasa corporal se ha
encontrado otro tipo de célula madre, denominada
mesenquimal que puede diferenciarse en numerosos tipos de
células de los tres derivados embrionarios
(musculares, vasculares, nerviosas, hematopoyéticas,
óseas). Aunque aún no se ha podido determinar
su relevancia fisiológica se están realizando
abundantes ensayos clínicos para sustituir tejidos
dañados (corazón) por derivados de estas
células.
La célula madre por excelencia es el cigoto,
formado cuando un óvulo es fecundado por un
espermatozoide. El cigoto es totipotente, es decir, puede dar
lugar a todas las células del feto y a la parte
embrionaria de la placenta.
Conforme el embrión se va desarrollando, sus
células van perdiendo esta propiedad (totipotencia) de
forma progresiva, llegando a la fase de blástula o
blastocisto en la que contiene células pluripotentes
(células madre embrionarias) capaces de diferenciarse en
cualquier célula del organismo salvo las de la parte
embrionaria de la placenta. Conforme avanza el desarrollo
embrionario se forman diferentes poblaciones de células
madre con una potencialidad de regenerar tejidos cada vez
más restringida y que en la edad adulta se encuentran en
"nichos" en algunos tejidos del organismo.

Meiosis
Meiosis es una de las formas de la reproducción
celular. Este proceso se realiza en las glándulas sexuales
para la producción de gametos. Es un proceso de
división celular en el cual una célula diploide
(2n) experimenta dos divisiones sucesivas, con la capacidad de
generar cuatro células haploides (n). En los organismos
con reproduccion sexual tiene importancia ya que es el mecanismo
por el que se producen los óvulos y espermatozoides
(gametos).1 Este proceso se lleva a cabo en dos divisiones
nucleares y citoplasmáticas, llamadas primera y segunda
división meiótica o simplemente meiosis I y meiosis
II. Ambas comprenden profase, metafase, anafase y
telofase.
Proceso celular
Los pasos preparatorios que conducen a la meiosis son
idénticos en patrón y nombre a la interfase del
ciclo mitótico de la célula. La interfase se divide
en tres fases:
Fase G1: caracterizada por el aumento de
tamaño de la célula debido a la
fabricación acelerada de orgánulos,
proteínas y otras materias celulares.Fase S :se replica el material genético, es
decir, el ADN se replica dando origen a dos cadenas nuevas,
unidas por el centrómero. Los cromosomas, que hasta el
momento tenían una sola cromátida, ahora tienen
dos. Se replica el 98% del ADN, el 2% restante queda sin
replicar.Fase G2: la célula continúa aumentando
su biomasa.
Meiosis I
En meiosis 1, los cromosomas en una célula
diploide se dividen nuevamente. Este es el paso de la meiosis que
genera diversidad genética.
Meiosis. Se divide en dos etapas. Meiosis I o fase
reductiva: su principal característica es que el material
genético de las células hijas es la mitad (n) del
de las células progenitoras (2n). Meiosis II o fase
duplicativa: las células resultantes de esta etapa tiene
el mismo contenido genético que sus células
progenitoras (n).
Profase I
La Profase I de la primera división
meiótica es la etapa más compleja del proceso y a
su vez se divide en 5 subetapas, que son:
Leptoteno: La primera etapa de Profase I es la etapa
del leptoteno, durante la cual los cromosomas individuales
comienzan a condensar en filamentos largos dentro del
núcleo. Cada cromosoma tiene un elemento axial, un
armazón proteico que lo recorre a lo largo, y por el
cual se ancla a la envuelta nuclear. A lo largo de los
cromosomas van apareciendo unos pequeños
engrosamientos denominados cromómeros. La masa
cromática es 4c y es diploide 2n.Zigoteno: Los cromosomas homólogos comienzan
a acercarse hasta quedar recombinados en toda su longitud.
Esto se conoce como sinapsis (unión) y el complejo
resultante se conoce como bivalente o tétrada (nombre
que prefieren los citogenetistas), donde los cromosomas
homólogos (paterno y materno) se aparean,
asociándose así cromátidas
homólogas. Producto de la sinapsis, se forma el
complejo sinaptonémico (estructura observable solo con
el microscopio electrónico).La disposición de
los cromómeros a lo largo del cromosoma parece estar
determinado genéticamente. Tal es así que
incluso se utiliza la disposición de estos
cromómeros para poder distinguir cada cromosoma
durante la profase I meiótica. Además el eje
proteico central pasa a formar los elementos laterales del
complejo sinaptonémico, una estructura proteica con
forma de escalera formada por dos elementos laterales y uno
central que se van cerrando a modo de cremallera y que
garantiza el perfecto apareamiento entre homólogos. En
el apareamiento entre homólogos también
está implicada la secuencia de genes de cada
cromosoma, lo cual evita el apareamiento entre cromosomas no
homólogos. Durante el zigoteno concluye la
replicación del ADN (2% restante) que recibe el nombre
de zig-ADN.Paquiteno: Una vez que los cromosomas
homólogos están perfectamente apareados
formando estructuras que se denominan bivalentes se produce
el fenómeno de entrecruzamiento cromosómico
(crossing-over) en el cual las cromátidas
homólogas no hermanas intercambian material
genético. La recombinación genética
resultante hace aumentar en gran medida la variación
genética entre la descendencia de progenitores que se
reproducen por vía sexual. La recombinación
genética está mediada por la aparición
entre los dos homólogos de una estructura proteica de
90 nm de diámetro llamada nódulo de
recombinación. En él se encuentran las enzimas
que medían en el proceso de recombinación.
Durante esta fase se produce una pequeña
síntesis de ADN, que probablemente está
relacionada con fenómenos de reparación de ADN
ligados al proceso de recombinación.Diploteno: Los cromosomas continúan
condensándose hasta que se pueden comenzar a observar
las dos cromátidas de cada cromosoma. Además en
este momento se pueden observar los lugares del cromosoma
donde se ha producido la recombinación. Estas
estructuras en forma de X reciben el nombre quiasmas. Cada
quiasma se origina en un sitio de entrecruzamiento, lugar en
el que anteriormente se rompieron dos cromatidas
homólogas que intercambiaron material genético
y se reunieron. En este punto la meiosis puede sufrir una
pausa, como ocurre en el caso de la formación de los
óvulos humanos. Así, la línea germinal
de los óvulos humanos sufre esta pausa hacia el
séptimo mes del desarrollo embrionario y su proceso de
meiosis no continuará hasta alcanzar la madurez
sexual. A este estado de latencia se le denomina
dictioteno.Diacinesis Esta etapa apenas se distingue del
diplonema. Podemos observar los cromosomas algo más
condensados y los quiasmas. El final de la diacinesis y por
tanto de la profase I meiótica viene marcado por la
rotura de la membrana nuclear. Durante toda la profase I
continuó la síntesis de ARN en el
núcleo. Al final de la diacinesis cesa la
síntesis de ARN y desaparece el
nucléolo.
Anotaciones de la Profase I
La membrana nuclear desaparece. Un cinetocoro se forma
por cada cromosoma, no uno por cada cromátida, y los
cromosomas adosados a fibras del huso comienzan a moverse.
Algunas veces las tétradas son visibles al microscopio.
Las cromátidas hermanas continúan estrechamente
alineadas en toda su longitud, pero los cromosomas
homólogos ya no lo están y sus centrómeros y
cinetocoros se encuentran separados.
Metafase I
El huso cromático aparece totalmente
desarrollado, los cromosomas se sitúan en el plano
ecuatorial y unen sus centromeros a los filamentos del
huso.
Anafase I
Los quiasmas se separan de forma uniforme. Los
microtúbulos del huso se acortan en la región del
cinetocoro, con lo que se consigue remolcar los cromosomas
homólogos a lados opuestos de la célula, junto con
la ayuda de proteínas motoras. Ya que cada cromosoma
homólogo tiene solo un cinetocoro, se forma un juego
haploide (n) en cada lado. En la repartición de cromosomas
homólogos, para cada par, el cromosoma materno se dirige a
un polo y el paterno al contrario. Por tanto el número de
cromosomas maternos y paternos que haya a cada polo varía
al azar en cada meiosis. Por ejemplo, para el caso de una especie
2n = 4 puede ocurrir que un polo tenga dos cromosomas maternos y
el otro los dos paternos; o bien que cada polo tenga uno materno
y otro paterno.
Telofase I
Cada célula hija ahora tiene la mitad del
número de cromosomas pero cada cromosoma consiste en un
par de cromátidas. Los microtubulos que componen la red
del huso mitótico desaparece, y una membrana nuclear nueva
rodea cada sistema haploide. Los cromosomas se desenrollan
nuevamente dentro de la carioteca (membrana nuclear). Ocurre la
citocinesis (proceso paralelo en el que se separa la membrana
celular en las células animales o la formación de
esta en las células vegetales, finalizando con la
creación de dos células hijas). Después
suele ocurrir la intercinesis, parecido a una segunda interfase,
pero no es una interfase verdadera, ya que no ocurre ninguna
réplica del ADN. No es un proceso universal, ya que si no
ocurre las células pasan directamente a la metafase
II.
Meiosis II
La meiosis II es similar a la mitosis. Las cromatidas de
cada cromosoma ya no son idénticas en razón de la
recombinación. La meiosis II separa las cromatidas
produciendo dos células hijas, cada una con 23 cromosomas
(haploide), y cada cromosoma tiene solamente una
cromatida.
Profase II
Profase Temprana: Comienzan a desaparecer la
envoltura nuclear y el nucleolo. Se hacen evidentes largos
cuerpos filamentosos de cromatina, y comienzan a condensarse
como cromosomas visibles.Profase Tardía II: Los cromosomas
continúan acortándose y engrosándose. Se
forma el huso entre los centríolos, que se han
desplazado a los polos de la célula.
Metafase II
Las fibras del huso se unen a los cinetocóros de
los cromosomas. Éstos últimos se alinean a lo largo
del plano ecuatorial de la célula. La primera y segunda
metafase pueden distinguirse con facilidad, en la metafase I las
cromatides se disponen en haces de cuatro (tétrada) y en
la metafase II lo hacen en grupos de dos (como en la metafase
mitótica). Esto no es siempre tan evidente en las
células vivas.
Anafase II
Las cromátidas se separan en sus
centrómeros, y un juego de cromosomas se desplaza hacia
cada polo. Durante la Anafase II las cromatidas, unidas a fibras
del huso en sus cinetocóros, se separan y se desplazan a
polos opuestos, como lo hacen en la anafase mitótica. Como
en la mitosis, cada cromátida se denomina ahora
cromosoma.
Telofase II
En la telofase II hay un miembro de cada par homologo en
cada polo. Cada uno es un cromosoma no duplicado. Se reensamblan
las envolturas nucleares, desaparece el huso acromático,
los cromosomas se alargan en forma gradual para formar hilos de
cromatina, y ocurre la citocinesis. Los acontecimientos de la
profase se invierten al formarse de nuevo los nucleolos, y la
división celular se completa cuando la citocinesis ha
producidos dos células hijas. Las dos divisiones sucesivas
producen cuatro núcleos haploide, cada uno con un
cromosoma de cada tipo. Cada célula resultante haploide
tiene una combinación de genes distinta. Esta
variación genética tiene dos fuentes: 1.- Durante
la meiosis, los cromosomas maternos y paternos se barajan, de
modo que cada uno de cada par se distribuye al azar en los polos
de la anafase I. 2.- Se intercambian segmentos de ADN.


Adaptaciones
generales de las células, muerte celular
a. Adaptaciones
celulares
Hiperplasia: es el aumento en el número de
células en un órgano o tejido, dando lugar
habitualmente a un aumento del volumen del órgano o
tejido. frecuentemente está asociada a la hipertrofia.
por ejemplo, el crecimiento inducido hormonalmente en el
útero implica a la vez un número aumentado de
células musculares y células epiteliales y el
agrandamiento de esas células. hiperplasia
fisiológica, la cual a su vez puede ser hormonal, que
aumenta la capacidad funcional de un tejido cuando se
necesita, por ejemplo, la proliferación del epitelio
glandular de la mama femenina en la pubertad y durante el
embarazo y, compensadora, que aumenta la masa tisular tras el
daño resección parcial, ejemplo, la
proliferación de las células hepáticas
residuales y la regeneración del hígado
después de una hepatectomía parcial. La
hiperplasia generalmente se debe a una producción
local aumentada de factores de crecimiento, niveles
aumentados de receptores de fc en las células
respondedoras, o activación de una determinada
vía de señalización intracelular. todos
estos cambios dan lugar a la producción de factores de
transcripción que activan muchos genes celulares,
incluyendo genes que codifican fc, de una
proliferación celular. En la hiperplasia hormonal, las
propias hormonas actúan como factores de crecimiento y
desencadenan la transcripción de diversos genes
celulares. el aumento de la masa tisular también se
consigue por el desarrollo de nuevas células a partir
de celulas madre. por ejemplo, la hiperplasia hepática
en la hepatitis crónica, en la cual está
comprometida la capacidad proliferativa de los hepatocitos.
hiperplasia patologica, la mayoría de las cuales
están producidas por un estímulo hormonal
excesivo o factores de crecimiento que actúan sobre
las células diana. por ejemplo, la hiperplasia
endometrial, el equilibrio entre estrógenos y
progesterona está alterado, lo cual da lugar a
aumentos absolutos o relativos de estrógenos con la
hiperplasia subsiguiente de las glándulas
endometriales y es causa frecuente de hemorragia menstrual
anormal. La hiperplasia prostática es otro ejemplo de
hiperplasia patológica en respuesta a los
andrógenos. La hiperplasia patológica
constituye un terreno fértil en el cual puede surgir
posteriormente la proliferación cancerosa. La
hiperplasia también es una respuesta importante de las
células del tejido conectivo en la curación de
heridas en las que, la proliferación de fibroblastos y
vasos sanguíneos ayuda a la reparación debido a
los factores de crecimiento, los cuales también
están implicados con ciertas infecciones
víricas, tales como el papiloma virus que produce
verrugas en la piel.Hipertrofia: se refiere a un aumento en el
tamaño de las células, lo que da lugar a un
aumento en el tamaño del órgano. el aumento del
tamaño de las células se debe a la
síntesis de más componentes estructurales
producida por un aumento de la demanda funcional o por
estimulación hormonal específica. por ejemplo,
los músculos de los físicoculturistas, son el
resultado de un aumento del tamaño de las fibras
musculares individuales en respuesta de la demanda.
así, la carga de trabajo está compartida por
una mayor masa de componentes celulares y a cada fibra
muscular se le ahorra un exceso de trabajo y de esta manera
escapa a la lesión. la célula muscular
agrandada consigue un nuevo equilibrio, permitiéndole
funcionar a un mayor nivel de actividad. El mecanismo implica
muchas vías de transducción de señal,
dando lugar a la inducción de un número de
genes que, a su vez, estimulan la síntesis de
numerosas proteínas celulares. en el corazón
existen dos grupos de señales: los desencadenantes
mecánicos tales como la distensión, y los
desencadenantes tróficos, tales como los factores de
crecimiento polipeptídicos y los agentes vasoactivos.
Finalmente alcanza un límite más allá
del cual el agrandamiento de la masa muscular ya no es capaz
de compensar el aumento de la carga y se sigue de
insuficiencia cardiaca. o sea que la hipertrofia puede ser
fisiológica o patológica.Atrofia: la disminución en el tamaño
de la célula por pérdida de sustancia celular
se conoce como atrofia y puede culminar con la muerte
celular. La atrofia fisiológica es común
durante el principio del desarrollo. Algunas estructuras
embrionarias, tales como la notocorda y el conducto tirogloso
sufren atrofia durante el desarrollo fetal. el útero
disminuye de tamaño poco después del parto. La
atrofia patológica depende de la causa subyacente y
puede ser localizada o generalizada: carga de trabajo
disminuida (atrofia por desuso), cuando un miembro roto se
inmoviliza con una escayola o cuando el paciente está
sujeto a un reposo completo en cama, rápidamente se
sigue de una atrofia muscular esquelética.
b. Muerte celular
La muerte celular programada o apoptosis es el conjunto
de reacciones bioquímicas que ocurren en las
células cuando se diferencian y ejercen funciones
normales, concluyendo tras un cierto número de divisiones
celulares con la muerte celular de una forma ordenada y
silenciosa; por lo que a la apoptosis se le conoce como muerte
celular programada.
En contraste con la necrosis, que es una forma de muerte
celular resultante de un daño agudo a los tejidos, la
apoptosis es un proceso ordenado, que generalmente confiere
ventajas al conjunto del organismo durante su ciclo normal de
vida. Por ejemplo, la diferenciación de los dedos humanos
durante el desarrollo embrionario requiere que las células
de las membranas intermedias inicien un proceso apoptótico
para que los dedos puedan separarse
La necrosis: de una célula sucede cuando
algún agente externo (traumatismo, tóxico,
agentes infecciosos, etc.) actúa sobre ella induciendo
su muerte. Las células que degeneran ocasionan una
serie de reacciones locales que conducen a respuestas de tipo
inflamatorio que son probablemente la manifestación
más importante de este proceso. La acción del
agente inductor de la necrosis produce una alteración
en las membranas plasmática y mitocondrial, donde se
alojan las bombas iónicas (fundamentalmente de Na+, K+
y Ca++) que se encargan de mantener el adecuado equilibrio
iónico intra-extracelular. Esta alteración en
los sistemas homeostáticos dispara un mecanismo de
defensa frente a la alteración de la homeostasis.
Así, el núcleo de la célula comienza a
transcribir ADN con información para la
síntesis de proteínas protectoras de la
célula (hsp – heat-shock proteins-, chaperonas).
En ocasiones estas proteínas son capaces de restaurar
las funciones celulares, pero en otras no y es entonces
cuando la célula continuará de manera
inevitable hacia su destrucción.Apoptosis: Desde el punto de vista del
envejecimiento, el proceso de eliminación de
células por apoptosis tiene una significación
funcional más importante. Durante el desarrollo
embrionario y en las fases posteriores además de la
proliferación celular se produce, de forma
fisiológica, un proceso de remodelación de los
órganos de la economía que implica la muerte
"programada" de numerosas células. Sin este proceso de
muerte celular programada nuestro organismo tendría
una morfología difícil de reconocer y
probablemente muchas de sus funciones estarían
comprometidas. Por medio de la MCP se eliminan células
que después de haber cumplido sus funciones,
fundamentalmente en el desarrollo, deben ser eliminadas. El
núcleo cambia notablemente de forma y se aprecia como
la cromatina, que normalmente está en forma de
eucromatina o cromatina dispersa (indica actividad
transcripcional del ADN), comienza a concentrarse formando
cromatina condensada o heterocromatina (indica que el ADN no
está transcribiendo). Finalmente todo el núcleo
se hace muy denso por la condensación total de la
cromatina (falta total de actividad transcripcional del ADN).
La consecuencia última de este proceso es la falta de
síntesis de ARN mensajero, ribosómico y de
transferencia, la imposibilidad de la síntesis de
proteínas y la consiguiente muerte y
fragmentación de la célula. Este proceso de
fragmentación se manifiesta morfológicamente
por la aparición de diferentes vesículas
esféricas (cuerpos apoptóticos), rodeados de
membrana celular, que contienen diversos organoides
citoplasmáticos degenerados. Estos cuerpos
apoptóticos van siendo fagocitados por los
macrófagos sin ningún tipo de reacción
inflamatoria acompañante. Esta ausencia de reacciones
locales del tipo inflamatorio es fundamental para entender la
"limpieza" biológica del proceso de apoptosis en
contraposición al proceso de necrosis.
Conclusión
Pese a la gran diversidad de seres vivos que existe
en la biosfera, todos ellos se caracterizan por estar
formados por unas pequeñas unidades llamadas
células.La célula es la base de la vida, tanto a
nivel estructural como a nivel funcional, ya que es la unidad
mínima de un organismo capaz de actuar de manera
autónoma.Existe gran diversidad en cuanto a la forma y el
tamaño de las células, pero todas tienen en
común:
-Membrana plasmática, es una estructura fina y
elástica que envuelve a la célula, la delimita, la
aísla del exterior pero permite el intercambio de
sustancias entre el medio interno y externo.
– Citoplasma, es el espacio interior delimitado por la
membrana plasmática en el que se encuentran los
orgánulos celulares.
-ADN, todas las células poseen una
molécula llamada ADN (ácido desoxirribonucleico).
Esta molécula contiene toda la información
genética del organismo y regula todos los procesos que
tienen lugar en el interior de la célula. Cuando el ADN
está libre en el citoplasma hablamos de células
procariotas. Cuando el ADN está incluido dentro de otra
membrana o envoltura llamada núcleo, hablamos de
células eucariotas.
Existen dos tipos de células eucariotas, que
se diferencian en:
– Célula vegetal: se caracterizan por tener una
pared celular rígida de celulosa que rodea la membrana
plasmática y le confiere resistencia a la célula;
poseen unas estructuras pigmentadas llamadas cloroplastos donde
se encuentran las clorofilas y poseen unas vacuolas de gran
tamaño donde almacenan sustancias de reserva.
–Célula animal: carecen de pared celular, por lo
que no tienen una forma definida; también carecen de
cloroplastos. Pueden tener vacuolas pero de tamaño mucho
menor que las de las células vegetales. Sin embargo tienen
centriolos, una estructura implicada en la división
celular de la que carecen las células
vegetales.
Bibliografía
Mayor, S., Pagano,
R.E . Pathways of clathrin-independent
endocytosis. 2007. Nature reviews in molecular and cell
biology. 8:603-612.Histología texto y atlas
color con biología celular y molecular.
Ff
Autor:
Leonardo Velasco
Prof. Dr. Victor Hugo
Cátedra: Histologia
1er Semestre T.S.U.
Citotecnología
San Cristóbal, Noviembre de
2012.
REPÚBLICA BOLIVARIANA DE
VENEZUELA
MINISTERIO DEL PODER POPULAR PARA LA
EDUCACIÓN SUPERIOR
CONVENIO UCV – UNET
ESTADO TÁCHIRA- SAN
CRISTÓBAL
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |