 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
As técnicas de estatística multivariadada que permitem a quantificação da dissimilaridade pela distância genética, assim como análises de agrupamentos, constituem-se em instrumentos adequados para a avaliação de acessos em bancos de germoplasma, onde o número de genótipos é elevado e vários descritores são utilizados (Pereira, 1989; Cruz, 1990).
Este trabalho teve por objetivo estimar a diversidade genética, bem como discriminar os caracteres morfoagronômicos de maior importância na caracterização da variabilidade de acessos do gênero Abelmoschus. Para tanto, utilizou-se técnicas de análise estatística multivariada, quais sejam, as análises de agrupamento hierárquico do vizinho mais próximo e componentes principais, com base na distância Euclidiana média padronizada.
Para o estudo da divergência genética com base em marcadores morfológicos, utilizaram-se 39 acessos do gênero Abelmoschus, sendo quinze pertencentes à espécie A. esculentus, quinze A. caillei, sete A. moschatus, um A. ficulneus e um A. tetraphyllus. Estes acessos foram cedidos pela Estação Regional da Geórgia do Departamento de Agricultura dos Estados Unidos (USDA), os quais foram solicitados com a intervenção da Embrapa Recursos Genéticos e Biotecnologia, seguindo-se a legislação para importação de germoplasma. Atualmente, os acessos estão sendo mantidos pelo Banco de Germoplasma da UENF. Os genótipos incluem espécies cultivadas e silvestres de diversas origens (Tabela 1).
As plantas foram cultivadas na Unidade de Apoio a Pesquisa da UENF, em Campos dos Goytacazes, no periodo de agosto de 1998 a janeiro de 1999. O espaçamento utilizado foi 1,0 m entre fileiras e 0,5 m entre plantas. Utilizou-se o delineamento experimental em blocos ao acaso, com 4 repetições e 5 plantas por parcela. Na caracterização do germoplasma utilizou-se 13 descritores quantitativos: número de segmentos do epicálice, comprimento do epicálice (mm), largura do epicálice (mm), comprimento do pedúnculo (cm), peso de 100 sementes (g), número de segmentos do estigma, altura de planta (cm), número de internós, comprimento da folha (cm), largura da folha (cm), nó do primeiro florescimento, nós produzindo frutos na haste principal, comprimento do fruto (cm) e 14 descritores qualitativos: tipo de fruto (1:quinado, 2:intermediário, 3:roliço); hábito de crescimento (1:ereto, 2:intermediário, 3:prostrado); presença de ramificação (1:baixa, 2:abundante); pubescência do caule (1:altamente piloso, 2:medianamente piloso, 3:notável); cor do caule (1:verde, 2:verde com manchas vermelhas, 3:vermelho); cor da folha (1:verde, 2:verde com nervuras vermelhas, 3:vermelha) forma do epicálice (1:linear, 2:lanceolado, 3:triangular); persistência dos segmentos do epicálice [1:não persistente até sete dias do florescimento, 2:parcialmente persistente (após sete dias), 3:persistente]; cor da pétala (1:amarela, 2:branca); coloração vermelha na base da pétala (1:apenas internamente, 2:ambos os lados); posição do fruto na haste principal (1:ereto, 2:horizontal, 3:pendular); cor do fruto imaturo (1:verde, 2:verde-escuro, 3:verde com manchas vermelhas, 4:vermelho, 5:vermelho-escuro); pubescência do fruto (1:pelos macios, 2:ásperos, 3:espinhosos); tipo do estigma (1:aberto, 2:semi-aberto, 3:fechado).
A caracterização foi efetuada de acordo com a lista de descritores do "International Plant Genetic Resources Institute" (IPGRI) elaborada por Charrier (1984). A análise estatística foi efetuada com base na distância Euclidiana média padronizada e componentes principais, utilizando-se os recursos computacionais do Programa Genes I (Cruz, 1997).
A Distância Euclidiana Média Padronizada é expressa por:

onde n = número de variáveis avaliadas; e Xij = a observação no acesso i em referência à característica j.
Para a padronização utilizou-se a relação:
![]()
em que S (Xj) é o desvio-padrão dos dados do j-ésimo caráter (Cruz & Regazzi, 1994).
Na obtenção dos componentes principais, as seguintes propriedades foram consideradas:
a) Se Yij é um componente principal, então:
![]()
b) Se yij' é outro componente principal, então:

ou seja, os componentes são não correlacionados.
c) Entre os componentes principais, yi1 apresenta a maior variância, yi2 o segundo maior e, assim, sucessivamente.
As estimativas desses parâmetros são obtidas pela solução do sistema de equação:
(R - ljI) aj = 0, em que lj são as raízes características ou autovalores da matriz de correlações entre as variáveis originais ou de covariâncias entre variáveis padronizadas. Existem p autovalores correspondentes às variâncias de cada um dos p componentes principais; aj = vetor característico ou também chamado autovetor, representa o conjunto de transformações ortogonais nas quais as variáveis originais padronizadas devem ser multiplicadas, para produzir as variáveis transformadas; I é a matriz identidade de dimensão p x p; e R é matriz de correlações fenotípicas entre pares de variáveis originais.
A importância relativa de cada componente (Irj) foi avaliada pela porcentagem da variância total que ele explica, ou seja:

sendo: V(Yi1) + V(Yi2) + .... + V(Yip) = Traço de R.
Adotou-se o critério de descartar a variável de maior coeficiente em cada componente, com autovalor acumulado maior que 0,70, de acordo com as proposições de Jolliffer (1973) e Pereira (1989).
Posteriormente, adotou-se o método hierárquico do vizinho mais próximo para o agrupamento. No procedimento analítico, partindo-se da matriz de distâncias genéticas, procedeu-se a sucessivas identificações dos genótipos mais próximos, a partir do par mais semelhante, até que se estabeleceu um diagrama de árvore. A distância entre um acesso k e um grupo formado pelos acessos i e j, é dada por: D2(ij)k = min (D2ik; D2jk), em que D2(ij)k é a distância entre o grupo ij e o acesso k, e min (D2ik; D2jk) é a menor distância entre os grupos de acessos ik e jk (Cruz & Regazzi, 1994).
Com base nas médias dos dados morfológicos obtidas a partir dos acessos, obtiveram-se três matrizes de distâncias Euclidianas, que foram analisadas pelo método hierárquico do vizinho mais próximo e de componentes principais.
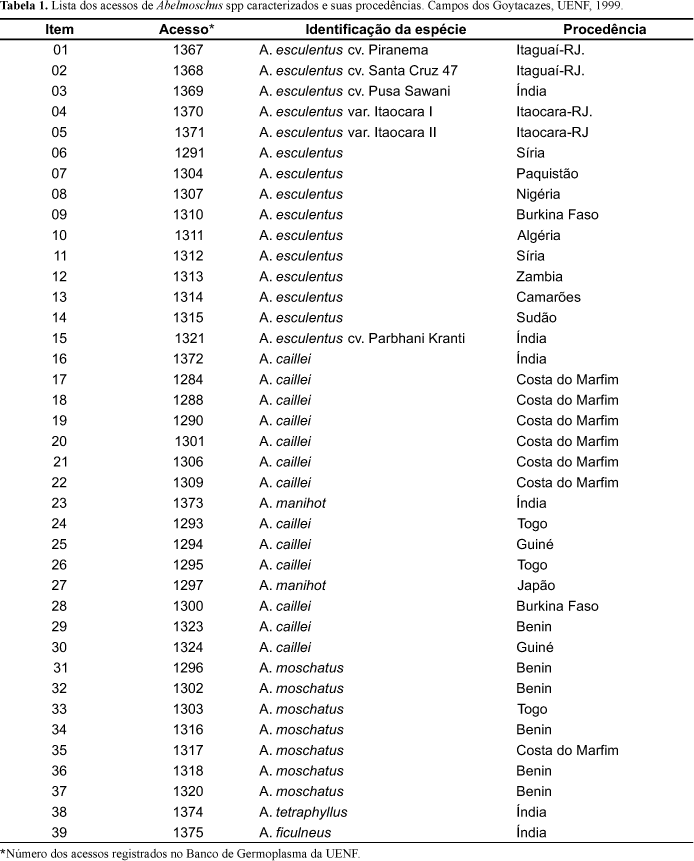
Com base na matriz de distâncias Euclidianas médias, geraram-se três dendrogramas (Figuras 1, 2, e 3). Fazendo-se um corte vertical a uma distância de ligação de 17,5 no agrupamento hierárquico dos acessos com base nas características avaliadas (Figura 1), observa-se que houve a formação de seis grupos. O grupo 1 reúne os acessos de A. esculentus e A. caillei. O segundo grupo reúne 5 dos 7 acessos de A. moschatus estudados. O grupo 3 reúne os acessos 1317 e 1320 de A. moschatus e os representantes de A. tetraphyllus e A. ficulneus. Os grupos 4, 5 e 6 foram compostos pelos acessos 1324 (A. caillei), 1297 (A. manihot) e 1373 (A. manihot), respectivamente.
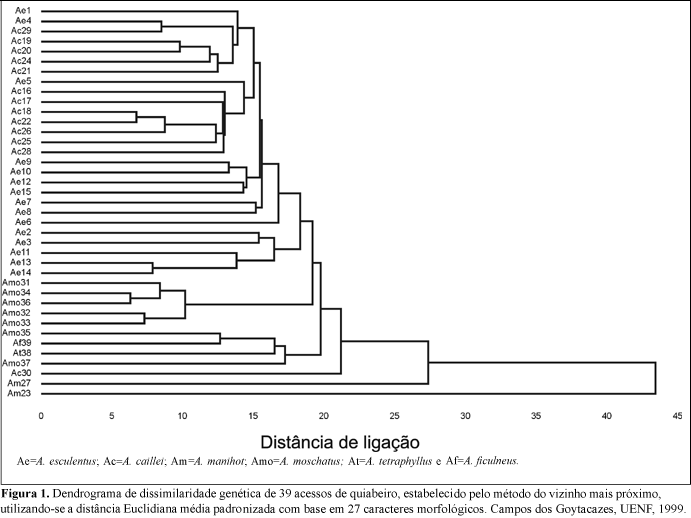
O método hierárquico agrupou os acessos de forma idêntica tanto para os 27 descritores (Figura 1) quanto para os 13 descritores quantitativos (Figura 2). Isto indica que os descritores qualitativos não afetaram o perfil da distribuição dos acessos no dendrograma, não interferindo, de forma determinante, no estudo da variabilidade entre genótipos de quiabeiro, com base em dados morfológicos. Os descritores quantitativos foram eficientes na discriminação dos acessos, porém, não foram capazes de classificar corretamente as espécies. Um dos motivos para isso, pode ser o fato de que estas características são controladas por muitos genes, sendo muito afetadas por fatores ambientais.
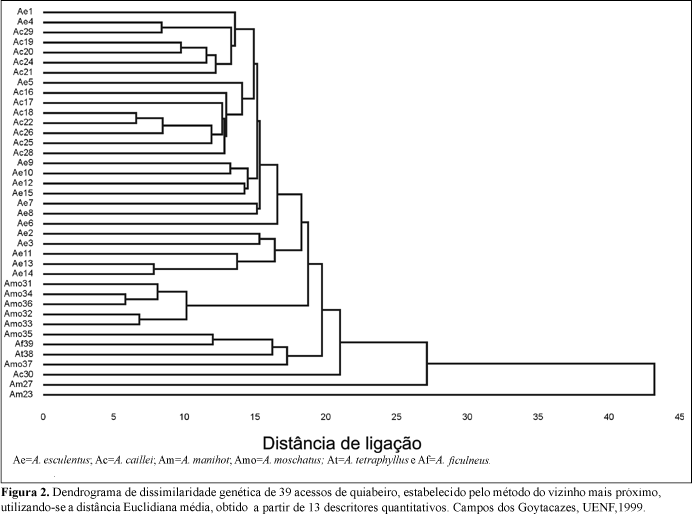
O agrupamento hierárquico, com base em características morfológicas qualitativas, reuniu os acessos conforme a espécie botânica, quando fez-se um corte vertical a uma distância de 2,5 (Figura 3). Isto ocorreu, provavelmente, pelo fato destas características serem controladas por poucos genes e, portanto, serem pouco afetadas pelo ambiente. Outrossim, é que a taxonomia clássica normalmente utiliza destas características para classificar as espécies. O dendrograma mostrou um baixo potencial em diferenciar os acessos dentro dos vários grupos formados, denotando uma reduzida variabilidade genética no germoplasma. Isto pode ser explicado pelo fato de que estas características, sendo controladas por poucos genes, não possibilitam um rastreamento muito abrangente dos genomas. Observou-se ainda, que não houve uma associação entre a diversidade genética e ecogeográfica entre os acessos.
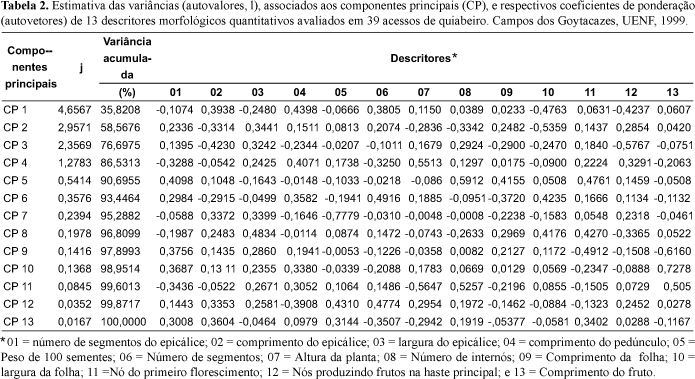
O resultado da análise de componentes principais, obtido a partir de 13 caracteres morfológicos quantitativos (Tabela 2), mostra que foram necessários quatro componentes, para que a variância por eles explicada atingisse um mínimo de 80%. Isso indica uma dispersão, de forma mais equitativa, da variância total, nos caracteres avaliados.
A técnica de análise multivariada de componentes principais tem a vantagem em relação aos métodos de análise univariada de avaliar a importância de cada característica estudada sobre a variação total disponível entre os acessos avaliados, possibilitando o descarte dos caracteres menos discriminantes, por já estarem correlacionados com outras variáveis redundantes ou pela sua invariância.
Com base nos resultados deste trabalho, foram descartados os caracteres: largura do epicálice, peso de 100 sementes, número de segmentos do estigma, altura da planta, comprimento da folha, largura da folha, nó do primeiro florescimento e comprimento do fruto. Porém o procedimento de descarte de variáveis pela técnica de componentes principais, pela possibilidade de eliminar variáveis que possuem pesos consideráveis nos primeiros componentes, principalmente quando estes acumulam grande fração da variância total (Morais, 1992).
A análise de componentes principais reduziu os treze descritores em três componentes principais, que explicaram 76,7% da variância total (Tabela 2). O componente 1, que explicou 35,8% da variância total, está associado a um contraste entre dois grupos de variáveis. Dentre as variáveis de maior peso neste componente pode-se citar: comprimento do epicálice, comprimento do pedúnculo, número de segmentos do estigma, largura da folha e nós produzindo frutos na haste principal. O componente 2, que explicou 22,7% está associado aos seguintes descritores: comprimento do epicálice, largura do epicálice, altura da planta, números de internós, largura da folha e nós produzindo frutos na haste principal. No componente 3, que sozinho explicou 18,1% da variância total, os descritores de maior peso foram: comprimento do epicálice, largura do epicálice, número de internós, comprimento da folha e nós produzindo frutos na haste principal.
A representação gráfica dos escores dos três primeiros componentes principais demonstra que houve a formação de quatro grupos (Figura 4). O primeiro e o maior deles é composto pelos acessos de A. esculentus e A. caillei, confirmando, desta forma, a aglomeração produzida pelos métodos já analisados. O segundo grupo é formado apenas pelos acessos de A. moschatus. O terceiro, reune três acessos de A. esculentus e apenas um de A. manihot. Por fim, o grupo quatro é composto pelos acessos de A. tetraphyllus e A. ficulneus, além de um acesso de A. manihot. Verifica-se, ainda, que ocorreu uma separação dos acessos de A. manihot. Estes acessos diferem, basicamente, pela variável altura da planta. Este fato não concorda com o método de agrupamento hierárquico com base nos descritores qualitativos, que revelaram os dois acessos de A. manihot como pertencentes ao mesmo grupo. Porém, está de acordo com os agrupamentos hierárquicos (Figuras 1 e 2).

Pode-se considerar que, de um modo geral, os dois métodos de aglomeração fundamentados em dados morfológicos, foram coerentes na formação dos grupos entre acessos de quiabeiro. Martinello et al. (1996), numa análise por componentes principais aplicada a características morfológicas em quiabeiro, verificaram que A. caillei foi a única das espécies silvestres que fez parte do grupo em que todos os outros acessos eram A. esculentus. Estes autores verificaram ainda que hibridações interespecíficas entre A. caillei (2n= 185-189) e A. esculentus (2n= 72-144), foram eficientes e que, os híbridos F1 se mostraram férteis e produziram grandes quantidades de sementes viáveis, apesar da diferença do número de cromossomos entre os parentais. Isto indica uma grande afinidade entre os genomas, que podem fazer parte de um mesmo complexo gênico. Estas constatações denotam que, no processo de evolução, estas espécies divergiram ou bifurcaram mais recentemente. Outra hipótese, proposta por Baum & Gupta (1996), sugere que A. caillei tenha se diferenciado a partir de A. esculentus, ou vice-versa, por um processo evolucionário reticulado, isto é, anfiploidização, poliploidização, hibridação ou outro sistema genético. Convém salientar que este tipo de situação envolvendo evolução reticulada para as plantas superiores, ainda é pouco estudada. Siemonsma (1982) hipotetizou que A. caillei é um anfiplóide natural de A. esculentus (2n= 130-140) e A. manihot (2n= 60-68), concordando com os resultados obtidos por esta pesquisa. Porém, para se entender plenamente como estas espécies evoluíram, deve-se recorrer às técnicas de citogenética molecular como, por exemplo, hibridização genômica in situ, uma vez que se trata de alopoliplóides.
Verificou-se ainda, que as técnicas estatísticas empregadas neste trabalho foram capazes de estabelecer padrões e classificações da diversidade genética, identificando claramente os grupos. As informações resultantes são úteis no entendimento da diversidade dentro da coleção e como está estruturada. As características morfométricas delineiam estes grupos auxiliando na pesquisa de novas cultivares e busca de espécies silvestres requeridas como base genética para programas de melhoramento da cultura.
ARIYO, O.J. Genetic diversity in West African okra (Abelmoschus caillei) (A. chev.) Stels - Multivariate analysis of morphological and agronomic characteristics. Genetic Resources and Crop Evolution, v. 40, p. 25-32, 1993.
BAUM, B.R.; GUPTA, P.K. Numerical taxonomy and genome analysis. In: JAUHAR, P.P., ed. Methods of genome analysis in plants. North Dakota: CRC Press, 1996. p. 345-356.
BISHT, I.S.; PATEL, D.P.; NAHAJAN, R.K. Classification of genetic diversity in Abelmoschus tuberculatus germplasm collection using morphometric data. Annual Applied Biology, v. 130, p. 325-335, 1997.
BOHAC, J.R.; AUSTIN, D.F.; JONES, A. Discovery of wild tetraploid sweetpotatoes. Economical Botany, v. 47, p. 193-201, 1993.
BRETTING, P.K.; WIDRLECHENER, M.P. Genetic markers and horticultural germplasm management. HortScience, v. 30, n. 7, p. 1349-1355, 1995.
BROWN, A.H.D.; CLEGG, M.T. Isozyme assessment of plant genetic resources. In: BATTAZI, M.C.; SCANDALIUS, J.G.; WHITT, G.S., eds. Isozymes: Current topics in biological and medical research. New York: A.R. Liss, 1983. p. 285-295.
CHARRIER, A. Genetic Resources of Abelmoschus (okra). Rome: IBPGR, 1984. 64 p.
CRAWFORD, D.J. 1990. Plant molecular systematics: macromolecular approaches. New York: Wiley, 1990. 388 p.
CRUZ, C.D. Programa Genes: aplicativo computacional em genética e estatística. Viçosa: Editora UFV, 1997. 394 p.
CRUZ, C.D.; REGAZZI, A.J. Modelos biométricos aplicados ao melhoramento genético. Viçosa: UFV - Imprensa Universitária, 1994. 390 p.
CRUZ, C.D. Aplicação de algumas técnicas multivariadas no melhoramento de plantas. Piracicaba: USP/ESALQ, 1990. 188 p. (Tese doutorado).
HAMON, S.; VAN SLOTEN, D.H. Characterization and evaluation of okra. In: BROWN, A.H.D.; FRANKEL, O; MARSHALL, D.R. ; WILLIAMS, J.T., eds. The use of plant genetic resources. Cambridge: Cambridge University Press, 1989. p. 173-196.
JOLLIFFER, I.T. Discarding variables in a principal component analysis, II. Real data. Applied Statistics, v. 22, p. 21-31, 1973.
MARTINELLO, G.E.; ABBOUD, A.C.S.; LEAL, N.R. Análise em componentes principais e de agrupamentos aplicada a características morfológicas e agronômicas em quiabeiro. Horticultura Brasileira, v. 14, n. 2, p. 200-203, 1996.
MORAIS, O.P. Análise multivariada da divergência genética dos progenitores, índices de seleção e seleção combinada numa população de arroz oriunda de intercruzamentos usando macho-esterilidade. Viçosa: UFV-Imprensa Universitária, 1992. 251 p. (Tese doutorado).
PEREIRA, A.V. Utilização de análise multivariada na caracterização de germoplasma de mandioca (Manihot esculenta Crantz). Piracicaba: USP/ESALQ, 1989. 108 p. (Tese doutorado).
SIEMONSMA, J.S. West-African okra morphological and cytogenetical indications for the existence of a natural amphidiploid of Abelmoschus esculentus (L.) Moench and A. manihot (L.) Medikus. Euphytica, v. 31, n. 1, p. 241-252, 1982.
Gilmar Efrem Martinello; Nilton R. Leal; Antônio T. Amaral Júnior; Messias G. Pereira; Rogério F. Daher - rogdaher[arroba]uenf.br
UENFCCTA, Av. Alberto Lamego, 2000, Horto, 28.015-620, Campos dos Goytacazes, RJ.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




