 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O trabalho foi executado em 12 tanques de alvenaria, sob um telado, no Centro Experimental do Instituto Agronômico de Campinas (IAC), Estado de São Paulo. O solo utilizado nas parcelas experimentais foi um Latossolo Roxo Distrófico (Oxissol, segundo Soil Taxonomy, 1990), cujas características físicas e químicas são apresentadas na Tabela 1.
Cada tanque medindo 2,0 m de comprimento por 0,5 m de largura por 0,4 m de profundidade, representou uma parcela. Esses tanques foram dispostos paralelamente entre si, distanciados de 0,5 m.
O solo de cada tanque foi adubado incorporando-se a uma profundidade média de 0,05 m, quantidades proporcionais a: 90 ppm de fósforo na forma de superfosfato simples; 40 ppm de potássio, na forma de cloreto de potássio e 30 ppm de nitrogênio, na forma de sulfato de amônio, efetuada no dia 11/05/1991.

Após 38 dias de emergência das plântulas em cada tanque procedeu-se a adubação de cobertura adicionando-se quantidade proporcional a 40 ppm de nitrogênio, na forma de sulfato de amônio. Aos 30 dias anteriores à semeadura o solo foi corrigido com aplicação de calcário proporcional à 3,0 t.ha-1.
A semeadura foi realizada no dia 21/05/91, em duas linhas espaçadas de 0,2 m, obtendo-se após a emergência (28/05/91) uma população proporcional de 180 plantas.m-2 (90 plantas/metro linear). Foi utilizada a cultivar IAC 287, material que apresenta o nome vulgar de YACO "S" e proveniente do cruzamento entre HIM (Heima=Frances)/COC (Cocoraque=75-S=Mexicano)/ /NAC(Nacozari 76=Mexicano), cujo ciclo é de aproximadamente 105 dias, da emergência à maturação fisiológica.
Foram efetuados dois tratamentos: T1 - controle, sem deficiência hídrica durante o ciclo da cultura mantendo-se o solo próximo a sua capacidade máxima de armazenamento de água através de irrigações efetuadas em dias alternados. T2 - deficiência hídrica, imposta pela suspensão da irrigação entre o 1º e o 13º dia após a floração, correspondentes a 71 e 83 dias após a emergência das plantas (DAE). A partir do 14º dia após a floração (DAF), correspondente a 84 DAE, foram realizados turnos de rega, em dias alternados com a finalidade de reconduzir o solo a uma umidade próxima a sua capacidade máxima de armazenamento. Os níveis de armazenamento foram determinados experimentalmente para cada parcela durante 45 dias anteriores à implantação do experimento, definindo-se como valor máximo para os tanques, um nível de umidade acima de 30% (massa/massa) e valor mínimo com 19% (massa/massa).
Durante o período de suspensão e reinicio da irrigação, foram monitorados durante 23 dias a umidade do solo (f), o potencial de água na folha bandeira (Yf), a taxa de fotossíntese da folha bandeira (tf), a condutância foliar (cf), o conteúdo relativo de água da folha (CRA), a variação da radiação fotossinteticamente ativa (RFA), a temperatura do ar (T) e a umidade relativa do ar (UR). A umidade do solo foi medida pelo método gravimétrico com amostras de solo coletadas com trado até profundidade de 0,2 m. O potencial de água na folha bandeira (Yf) foi medido com higrômetros (C-30, Wescor, Inc.) acoplados a um microvoltímetro de ponto de orvalho (HR 33T, Wescor). As 8 e 14 horas amostras de 2,0 cm2 da parte mediana da folha bandeira foram colhidas e acomodadas diretamente nos higrômetros.
Após 3 h para entrada em equilíbrio determinou-se em laboratório o valor de potencial (Yf) pelo método do ponto de orvalho. Cada medida foi efetuada com seis repetições. Os higrômetros foram calibrados anteriormente com soluções de cloreto de sódio. A taxa de fotossíntese foi determinada em um sistema fechado com um aparelho portátil para medida da fotossíntese (LI-6200, LICOR) com câmara de 0,25 L, no período diário compreendido entre 8 e 15 h, em intervalos de uma hora. O sensor do analisador foi sempre colocado em posição intermediária da folha bandeira em plantas marcadas no início do período reprodutivo para as referidas determinações. A umidade relativa do ar na câmara de fotossíntese foi mantida em valores aproximadamente iguais aos do ar ambiente. A condutância foliar, a transpiração, a temperatura da folha e a radiação fotossinteticamente ativa foram também medidas com o mesmo equipamento (LI-6200). A determinação do conteúdo relativo de água das folhas (CRA) foi realizado às 8 e 14 horas de cada dia, em dois grupos distintos de amostragens, correspondentes aos tratamentos estudados. Para análise foram utilizadas as folhas bandeiras, através do seguinte procedimento: após o corte as folhas foram rapidamente colocadas em vidros hermeticamente fechados e com seu interior envolto em papel filtro umedecido. Em laboratório foram coletados discos das folhas com 0,9 cm de diâmetro utilizando-se de um vasador. Os discos foram pesados em balança analítica de precisão, na condição de massa verde, após três horas em água destilada (massa túrgida) e após 48 horas em estufa de ventilação forçada à 60°C (massa seca). Os valores obtidos nestas pesagens foram utilizados na seguinte expressão (WERTHERLEY, 1950), para se obter o CRA (%):
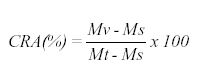
onde, Mt é a massa túrgida, Ms a massa seca e Mv a massa verde.
O material para análise da fitomassa foi coletado após a floração (07/08/91) em três oportunidades, possibilitando a medida das massas secas dos colmos mais bainhas, das folhas verdes e senescentes e das panículas. Cada coleta constou de 60 plantas contidas nos 0,2 m de linha dupla (30 plantas/linha), sendo a massa seca medida após secagem do material em estufa de circulação forçada de ar à 70° C por 72 horas. Na última coleta em vinte espigas por repetição determinaram-se os componentes de produtividade (número de espiguetas/espiga, número de espigas/planta, número de grãos/espiga e a massa seca de 500 grãos) e o índice de colheita considerado no caso como a relação entre a massa seca de grãos pela massa seca total multiplicado por 100.
O delineamento experimental foi blocos ao acaso com parcelas subdivididas e seis repetições, sendo as parcelas principais representadas pelos tratamentos e as sub-parcelas pelas coletas.
A suspensão da irrigação no tratamento com deficiência hídrica provocou uma queda do potencial da água na folha (Yf) em relação às plantas do tratamento sem deficiência hídrica (Figura 1). A queda do potencial de água nas folhas das plantas sem deficiência hídrica deve-se certamente à tendência natural de diminuição com a idade da folha.
No tratamento com deficiência hídrica o solo atingiu o nível crítico de 19% de umidade (0,19 kg de água/kg de solo) após o 11º dia sem irrigação, sendo que o efeito visual nas plantas ocorreu após 48 horas. Os sintomas de deficiência hídrica nas plantas começaram a aparecer, quando o Yf atingiu valor de -1,8 Mpa, correspondente à sete dias após floração. No 13º dia após a floração este tratamento atingiu o valor mínimo de -3,1 Mpa, quando então foi realizada irrigação até o solo atingir a umidade de 30% (0,3 kg de água/kg de solo). Portanto neste tratamento a deficiência hídrica ocorreu desde o 71 DAE, prolongando-se por 15 dias do desenvolvimento dos grãos, sendo mais acentuada aos 83 dias após emergência. Comparativamente, o Yf às 14 h atingiu valores menores que às 8 h, em função da maior demanda respiratória e o comportamento diário da temperatura do ar. O potencial de água da folha no tratamento com deficiência às 14 h recuperou-se parcialmente após seis dias do reinicio da irrigação, porém não apresentando uma recuperação total (Figura 1), devido basicamente ao efeito drástico do estresse hídrico e a idade avançada das folhas.

Figura 1. Potencial da água na folha bandeira do trigo cv. IAC-287 submetido a deficiéncia hídrica em diferentes fases durante o periodo reprodutivo; sem deficiencia hídrica determinaçôes 87 é 14 horas; con irrigaçâo suspensa entre o 1º e o 13º dia após floraçâo (DAF), determinaçôes 8 horas e 14 horas.
As curvas de fotossíntese nas plantas em condições naturais sem deficiência hídrica, independente- mente da idade da folha bandeira, descritos na literatura para o trigo e outras espécies com metabolismo fotossintético do tipo C3 (MACHADO et al., 1993) apresentam a forma de uma hipérbole regular. No presente caso são apresentados os incrementos de fotossíntese, em observações realizadas das 8 às 15 horas, no tratamento com deficiência hídrica, desconsiderando o decaimento diário da fotossíntese e irradiância, o que ocorre mesmo em dias considerados límpidos (Figura 2). Tal comportamento está de acordo com os obtidos para outras cultivares de trigo ( RAWSON et al., 1983). Porém, tanto o valor máximo da fotossíntese como a irradiância em que ocorreu a saturação lumínica, decresceu com a idade da folha, não havendo recuperação nos dias 14, 15, 16 e 17 após a floração e permanecendo num patamar baixo e quase constante no final do ciclo, dia 24.
A taxa de fotossíntese aumentou com a irradiância, cujos valores mais elevados ocorreram em horários próximos às 13 h, quando as temperaturas foram mais elevadas e a umidade relativa do ar menor (Figura 3), tendo como conseqüência, maior demanda transpiratória das plantas. Porém, sob deficiência hídrica a demanda transpiratória não é atendida e conseqüentemente, ocorre o fechamento dos estômatos e queda da fotossíntese (MACHADO et al., 1993).
Na Figura 2 verifica-se que a partir do primeiro dia de suspensão da irrigação foi obtido em média um valor de 24,9 mmol.m-2.s-1, cujos valores decresceram para 23,8 aos 7 DAF; 21,2 aos 10 DAF e 14,6 mmol.m-2.s-1 aos 13 DAF, considerado o dia crítico em termos de deficiência hídrica. Os incrementos da taxa fotossintética diminuíram à medida que aumentou a deficiência hídrica, sendo respectivamente, de 12,6 mmol.m-2.s-1 no primeiro DAF e 4,5 mmol.m-2.s-1 aos 13 DAF.
Os valores máximos da taxa fotossintética apresentados foram: 14,3 mmol.m-2.s-1 aos 14 DAF; 11,2 mmol.m-2.s-1 aos 15 DAF; 12,8 mmol.m-2.s-1 a os 16 DAF e 12,9 mmol.m-2.s-1 aos 17 DAF.
Interessante observar que, mesmo reiniciando a irrigação aos 14 DAF o efeito mais drástico da deficiência hídrica na planta se manifestou somente aos 16 DAF, para posteriormente apresentar uma tendência de recuperação, o que pode ser notado pelos respectivos incrementos diários da taxa de fotossíntese, de 4,5 mmol.m-2.s-1 aos 14 DAF; 3,4 mmol.m-2.s-1 aos 15 DAF; 3,0 mmol.m-2.s-1 aos 16 DAF e 3,0 mmol.m-2.s-1 aos 17 DAF. Nos demais dias ocorreu comportamento similar na taxa de fotossíntese com valores máximos de 13,5 mmol.m-2.s-1 aos 18 DAF; 14,1 mmol.m-2.s-1 aos 20 DAF; 14,1 mmol.m-2.s-1 aos 23 DAF e 14,2 mmol.m-2.s-1 aos 24 DAF.


Porém, os valores do incremento diário da taxa de fotossíntese se mantiveram próximos dos valores de 5,7 mmol.m-2.s-1 aos 18 DAF; 6,2 mmol.m-2.s-1 aos 20 DAF; 6,2 mmol.m-2.s-1 aos 23 DAF e 6,3 mmol.m-2.s-1 aos 24 DAF. Comparativamente, as taxas de fotossíntese diária apresentaram uma tendência a aumentar, a partir de irradiância acima de 670 mmol.m-2.s-1, relacionadas com o aumento da temperatura e diminuição da umidade relativa do ar (Figura 3). Apesar desta recuperação os efeitos da deficiência hídrica sobre o crescimento e pegamento dos grãos foram significativos. Após o reinicio da irrigação houve uma leve tendência de recuperação da fotossíntese. A maior variabilidade dos valores de fotossíntese podem estar relacionados com a aceleração da senescência das folhas e pode indicar heterogeneidade na recuperação destes órgãos. As diferenças nos níveis de recuperação da fotossíntese no tratamento indica sua dependência sobre intensidade e época de ocorrência da deficiência hídrica, como sugeriu SLAVIK (1975), ao comparar parcelas irrigadas com parcelas submetidas a deficiência hídrica na cultura do trigo (Figura 4). Em função da radiação fotossinteticamente ativa, verifica-se que o efeito negativo da deficiência hídrica pode advir da redução no processo de translocação de metabólitos e principalmente da taxa de fotossíntese. Nesta figura, são mostrados valores menores da taxa fotossintética para as parcelas estressadas em relação às parcelas irrigadas. Tal comportamento indica que sem deficiência hídrica a produção fotossintética excedeu a demanda dos grãos, sendo o excedente alocado para os colmos formando reservas como também foi observado por FISCHER (1983), MACHADO et al.(1993), MANFRON et al. (1996).
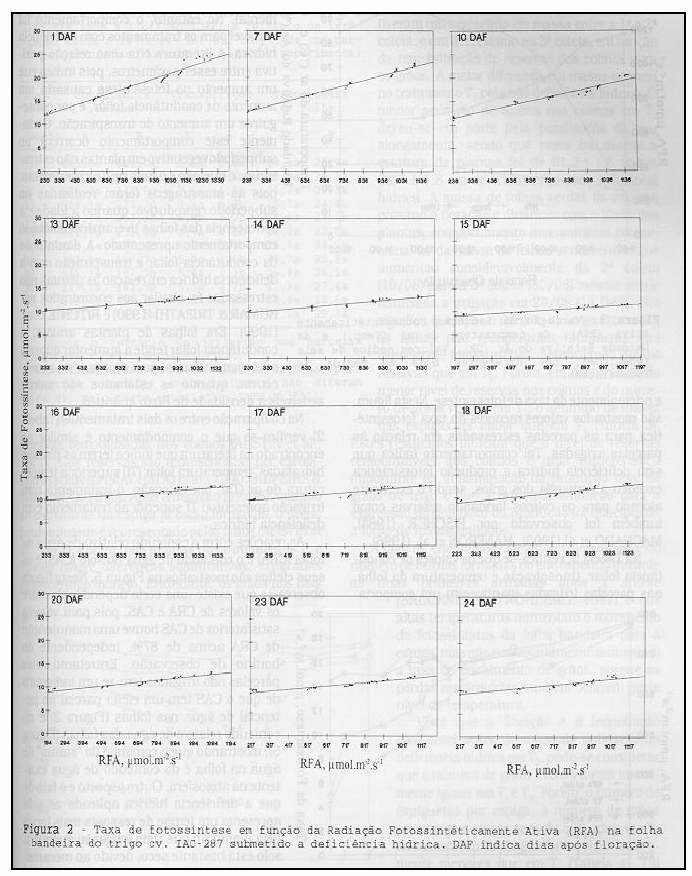
Na Tabela 2, verifica-se que os valores de condutância foliar, transpiração e temperatura da folha, nas parcelas irrigadas mantiveram um comportamento homogêneo, não apresentando diferença significativa durante o período experimental. No entanto, o comportamento foi variável para os tratamentos com deficiência hídrica. A literatura cita uma relação positiva entre esses parâmetros, pois indica que um aumento na fotossíntese causaria um aumento de condutância foliar, e por conseguinte um aumento de transpiração. Certamente este comportamento ocorrerá no subperíodo vegetativo em plantas não estressadas, o que não é o caso neste experimento, pois as amostragens foram realizadas no subperíodo reprodutivo, quando a idade e a senescência das folhas tiveram influência no comportamento apresentado. A diminuição da condutância foliar e transpiração com a deficiência hídrica em relação às plantas não estressadas foi similar aos encontrados por KUMAR & TRIPATHI (1990) e RITCHIE et al. (1990). Em folhas de plantas anuais, a condutância foliar tende a aumentar quando se tornam maturas, para diminuir na senescência, quando os estômatos são menos sensíveis à densidade de fluxo quântico.
Na comparação entre os dois tratamentos (Tabela 2) verifica-se que o comportamento é similar ao encontrado na literatura que indica terem as plantas hidratadas, temperatura foliar (Tf) superior a temperatura do ar (Ta), por extensão, o tratamento com irrigação apresentou Tf superior ao tratamento com deficiência hídrica.
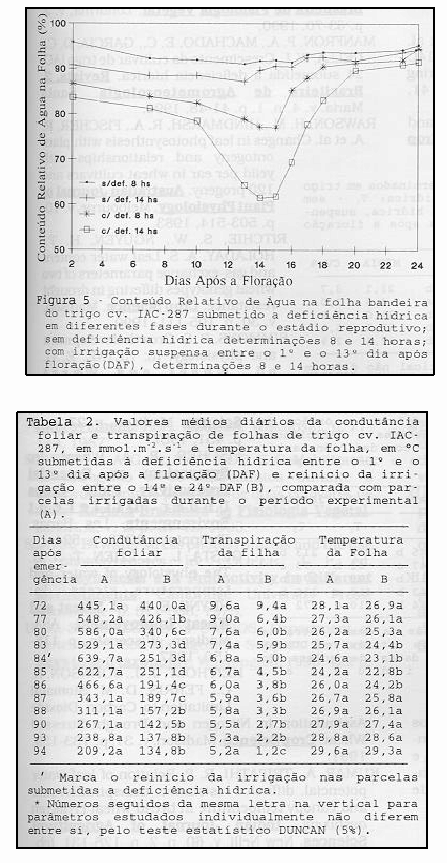
As relações entre o conteúdo relativo de água na folha (CRA) e o conteúdo de água no solo (CAS) e os seus efeitos são mostrados na Figura 5. Nesta figura observa-se que existe uma certa dependência entre os valores de CRA e CAS, pois para valores satisfatórios de CAS houve uma manutenção de CRA acima de 87%, independente do horário de observação. Entretanto, nas parcelas não irrigadas tem-se um indicativo de que o CAS tem um efeito parcial no potencial de água nas folhas (Figura 2) e no conteúdo relativo de água das folhas (Figura 5), mostrando que o comando do "status" de água na folha é do conteúdo de água existente na atmosfera. Outro aspecto é o fato de que a deficiência hídrica aplicada ao solo apresenta um tempo de resposta mais longo pela planta do que uma irrigação quando o solo está bastante seco, devido ao mecanismo de defesa da planta. As tendências observadas nos resultados deste trabalho são similares as encontradas por RITCHIE et al. (1990) e XU & ISHII (1990,1991), ao estudarem o comportamento de plantas de trigo submetidas a deficiência hídrica.
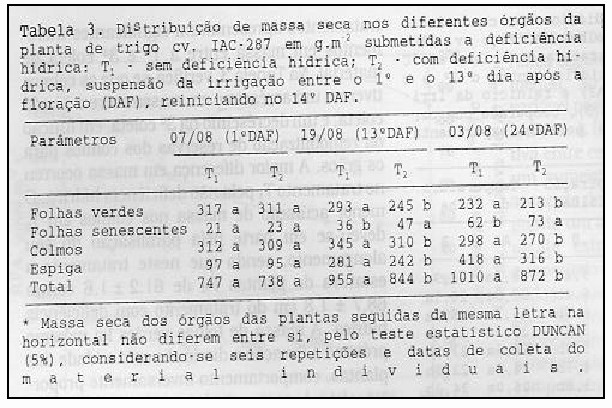
Em trigo cerca de 10 a 60% da massa acumulada nos grãos pode ser originada de reservas remobilizadas dos colmos (AUSTIN et al., 1977). Tais reservas são representadas por carboidratos solúveis (MACHADO et al., 1990; SILVEIRA & MACHADO, 1990). Após a floração (07/08), quando as estruturas da espiga já estão formadas, o dreno predominante é representado pelos grãos (AUSTIN et al., 1977). Porém, nas plantas dos tratamentos T1 e T2 não houve diferenças significativas, explicadas pelo idêntico manejo. Entretanto, os grãos nos dois tratamentos apresentaram significantes incrementos em massa entre a 1ª e 3ª coleta de material. Na Tabela 3, verifica-se que os colmos tiveram um acréscimo em massa entre a 1ª e 2ª coleta, e um decrésci mo na 3ª coleta, em função da remo bilização de reservas dos colmos para os grãos. A maior diferença em massa ocorreu no tratamento T1 pela não deficiência hídrica. O menor acúmulo de massa nos colmos em T2 deveu-se em parte pela paralisação do seu alongamento, sendo que neste tratamento a estatura de plantas foi de 61,2 ± 1,6 contra 68,7 ± 1,8 cm do tratamento com deficiência hídrica. A massa de folhas verdes foi máximo próximo a floração e diminuiu com a idade das plantas, comportamento inversamente proporcional ao da massa de folhas senescentes, que aumentou consideravelmente da 2ª coleta (19/08) para a 3ª coleta (30/08) mesmo sendo reiniciada a irrigação em 20/08 (14 DAF). Isto significa que após o reinicio da irrigação em T2, as folhas não recuperaram totalmente sua capacidade fotossintética, acumulando menos reservas que em T1. Porém, em decorrência do menor nível de reservas nos colmos e do número de grãos viáveis em T1, o acúmulo de massa nas espigas foi maior que em T2 (Tabela 3). Estes resultados indicam que o nível de reservas remobilizáveis dos colmos é uma característica importante na determinação da produção final de grãos de trigo e que a principal limitação para a expressão do potencial da produção foi relacionada à fonte de fotoassimilados.
A taxa de enchimento de grãos é dependente do número de células formadas no endosperma durante as duas primeiras semanas após a antese (BRUCKNER & FROHBERG, 1987) e que altas temperaturas aumentam o movimento de fotossintatos da folha bandeira para a espiga, mas não necessariamente aumentam a taxa de enchimento de grãos, porque as perdas respiratórias também ocorrem neste nível de temperatura.
Visto que a floração e a fecundação ocorreram antes do início da aplicação da deficiência hídrica em T2, pode-se considerar que o número de grãos viáveis foram inicialmente iguais em T1 e T2. Porém, o número de espiguetas por espiga, o número de grãos por espiga e massa seca de 500 grãos em T2 na maturação fisiológica foram significativamente menores que em T1 (Tabela 4). Tal resultado pode indicar que, mesmo após a ocorrência da fecundação, o número de grãos que completam seu desenvolvimento pode diminuir em conseqüência da ocorrência de deficiência hídrica. O índice de colheita maior no tratamento T1 em relação ao T2 é um indicativo de que a translocação de carboidratos de outras partes da planta para os grãos foi mais eficaz neste tratamento. Em decorrência de uma menor capacidade fotossintética em T2 e de sua lenta recuperação após o reinicio da irrigação, foram obtidos índices não satisfatórios de produtividade (Tabela 3).

A deficiência hídrica induz queda significativa das taxas de fotossíntese na cultivar de trigo IAC-287.
A susceptibilidade à deficiência hídrica não representa perda total de produção e apresenta um controle fisiológico eficiente na manutenção da turgescência das folhas.
AUSTIN, R. B., EDRICH, J. A., FORD, M.A. et al. The rate of the dry matter, carbohydrates and 14C lost from the leaves and stems of wheat during grain filling. Annals of Botany, Oxford, v. 41, p. 1309- 1321, 1977.
BRUCKNER, P. L., FROHBERG, R. G. Rate and Duration of Grain Fill in Spring Wheat. Crop Science, Madison, v. 27, n. 3, p. 451, 1987.
FISCHER, R. A. Wheat. In: Symposium on Potential Productivity Field Crops Under Different Environments, Las Banas, Philippines, 1983, p. 524.
GUSTA, L. V., CHEN, T. N. H. The physiology of water and temperature stress. In: HEYNE, E. G. ed. Wheat and Wheat Improvement. ASA. Madison, Wisconsin, p. 115-150, 1987.
HEITHOLT, J. J., JOHNSON, R. C., FERRIS, D. M. Stomatal Limitation to Carbon Dioxide Assimilation in Nitrogen and Drought-Stressed Wheat. Crop Science, Madison, v. 31, p. 133- 139, 1991.
KUMAR, A., TRIPATHI, R. P. Relation of leaf-water potencial, diffusice resistance, transpiration rate and canopy temperature in bread wheat (Triticum aestivum). Indian Journal of Agricultural Sciences, New Nelli, v. 60, n. 2, p. 128-131, feb. 1990.
MACHADO, E. C., LAGÔA, A. M. M. A., TICELLI, R. Relações Fonte-Dreno em Trigo Submetido a Deficiência Hídrica no Estádio Reprodutivo. Revista Brasileira de Fisiologia Vegetal. São Carlos, v. 5, n. 2, p. 145-150, 1993.
MACHADO, E. C., SILVEIRA, J. A. G. da, BASTOS, C. R. Trocas de CO2, acúmulo de fitomassa e remobilização de reservas durante o crescimento de panículas de duas cultivares de arroz. Revista Brasileira de Fisiologia Vegetal, Londrina, v. 2, p. 63-70, 1990.
MANFRON, P. A., MACHADO, E. C., GARCIA, D. C., CELLA, W. L. Crescimento da cultivar de trigo IAC-24 submetida a deficiência hídrica. Revista Brasileira de Agrometeorologia. Santa Maria, v. 4, n. 1, p. 41-48, 1996.
RAWSON, H. M., HINDMARSH, R. A., FISCHER, R. A. et al. Changes in leaf photosynthesis with plant ontogeny and relationships with yeild per ear in wheat cultivars and 120 progeny. Australian Journal of Plant Physiology, Melbourne, v. 10, p. 503-514, 1983.
RITCHIE, S. W., NGUYEN, H. T., HOLADAY, A. S. Leaf water content and gas-exchange parameters of two wheat genotypes differing in drought resistance. Crop Science, Madison, v. 30, p. 105-111, 1990.
SIMMONS, R. S. Growth development and physiology. In: HEYNE, E. G. ed. Wheat and Wheat Improvement. ASA. Madison, Wisconsin, p. 77-113, 1987.
SILVEIRA, J. A. G. da, MACHADO, E. C. Mobilização de nitrogênio e de carboidratos durante o desenvolvimentode panículas de duas cultivares de arroz. Revista Brasileira de Fisiologia Vegetal, Londrina, v. 2, p. 37-46, 1990.
SLAVIK, B. Water stress photosynthesis and the use of photosynthates. In: COOPER, J. P. ed. Photosynthesis and Productivity in Different Environments. Cambridge University Press, Cambridge, p. 511-536, 1975.
USDA-SMSS. ESTADOS UNIDOS. Department of Agriculture. Soil Management Support. Services. Keys to soil taxonomy. Washington, 1990. 422 p.
WERTHERLEY, P. E. Studies in the water relations of cotton plant. I-The field measurement of water deficits in leaves. New Phytologist, Cambridge, v. 49, p. 81-97, 1950.
XU, H., ISHII, R. Effects of water deficits on photosynthesis in wheat plants: V Difference among plant in water relations. Japanese Journal of Crop Science, Tokyo, v. 59, n. 2, p. 384-389, 1990.
XU, H., ISHII, R. Effects of water deficits on photosynthesis in wheat plants: VI Capacitance and resistance of transpiratory water flux in different plant parts. Japanese Journal of Crop Science, Tokyo, v. 6, n. 1, p. 75-81, 1991.
|
Revista Brasileira de Agrometeorologia, Santa Maria, v. 4, n. 2, p. 39-47, 1996. |
Paulo Augusto Manfron2, Eduardo Caruso Machado3, Danton Camacho Garcia4, Willyans Luiz Cella5
manfronp[arroba]smail.ufsm.br
1. Extraído da Tese de Doutorado do primeiro autor. ESALQ/USP: Programa de Pós-graduação em Agronomia, dezembro/92.
2. Engº Agrº, Doutor, Prof. Adjunto Departamento de Fitotecnia/CCR/UFSM. 97.119-900. Santa Maria/RS. Pesquisador do CNPq.
3. Engº Agrº, Doutor, Pesquisador Científico (PqC), Seção de Fisiologia, Instituto Agronômico de Campinas (IAC)/SP. Pesquisador do CNPq.
4. Engº Agrº, Mestre, Prof. Assistente Departamento de Fitotecnia/CCR/UFSM. 97.119-900. Santa Maria/RS.
5. Bolsista PIBIC/CNPq, aluno do Curso de Agronomia/CCR/UFSM. 97.119-900, Santa Maria/RS.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




