 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O experimento foi conduzido nas instalações do Setor de Avicultura do Departamento de Zootecnia do Centro de Ciências Agrárias da Universidade Federal de Viçosa (UFV) - MG no período de maio a agosto de 2003 (112 dias de duração).
Foram utilizadas 360 codornas fêmeas (Coturnix coturnix japonica) com 56 dias de idade e peso médio de 121,5 g durante quatro períodos de 28 dias.
As aves foram alojadas em gaiolas de arame galvanizado (1,0 m de comprimento x 0,23 m largura x 0,20 m altura) dispostas em três andares, montadas em esquema de escada. Cada gaiola foi subdividida em duas repartições de 0,50 m, com capacidade para dez aves por repartição, fornecendo área de 115 cm2/ave. Sobre o piso de cimento, logo abaixo das gaiolas, foi colocada uma camada de maravalha para absorção da umidade das excretas. O comedouro e o bebedouro eram do tipo calha, em chapa metálica galvanizada, e ambos foram colocados percorrendo toda a extensão das gaiolas, sendo o comedouro posicionado na parte frontal e o bebedouro na parte posterior das gaiolas.
O delineamento experimental foi inteiramente casualizado, em esquema fatorial 6 x 4, sendo constituído de seis tratamentos (níveis de Ca na dieta), cada um com seis repetições, e quatro períodos de 28 dias, sendo a unidade experimental representada por uma gaiola contendo dez aves.
Os tratamentos foram constituídos de seis dietas isocalóricas, isoprotéicas e isoaminoacídicas, à base de milho e farelo de soja, diferenciadas quanto ao nível de Ca (1,6; 2,0; 2,4; 2,8; 3,2 e 3,6%), conforme representado na Tabela 1. Essas dietas foram formuladas com base na composição dos ingredientes, descrita por Rostagno et al. (2000).
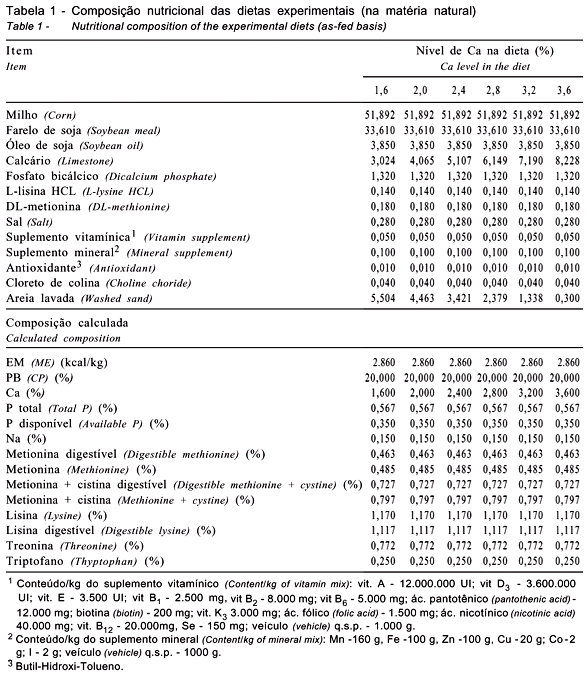
As exigências nutricionais utilizadas para a formulação das dietas das aves, exceto as de Ca, metionina+cistina (met+cis) e lisina, foram calculadas com base nas recomendações preconizadas pelo NRC (1994). As exigências de met+cis e lisina foram estimadas com base nas recomendações de Pinto (2003a,b), respectivamente, dando continuidade aos trabalhos de exigências nutricionais desenvolvidos na UFV com a espécie Coturnix coturnix japonica. A ração e a água foram fornecidas à vontade durante todo o período experimental. O programa de iluminação foi iniciado quando as aves atingiram 40 dias de idade, sendo estabelecidas inicialmente 14 horas de luz por dia, com aumentos semanais de 30 minutos de luz até atingir 17 horas de luz por dia, programa mantido até o término do período experimental. Este período de luz foi controlado por um relógio automático (timer) que permitia o acender das luzes à noite e na madrugada, conforme procedimento adotado nas granjas comerciais.
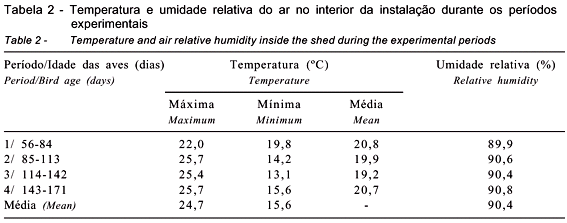
A temperatura e a umidade relativa no galpão foram monitoradas duas vezes ao dia por termômetros de mínima e máxima e de bulbos seco e úmido distribuídos em pontos estratégicos no local, posicionados à altura das aves. As médias de temperatura e de umidade obtidas durante o período experimental são descritas na Tabela 2.
Foram avaliados o desempenho produtivo, a qualidade dos ovos e o tecido ósseo das aves. Na avaliação do desempenho produtivo, foram considerados a produção média de ovos por ave/dia (%), a produção de ovos comerciais (%), o consumo de ração (g/ave/dia), o peso dos ovos (g), a massa de ovos (g/ave/dia), a conversão alimentar (g de ração por dúzia e por massa de ovos) e o ganho de peso corporal (g).
A qualidade dos ovos foi determinada por meio de análises físico-químicas da casca dos ovos. Nas análises físicas, foram incluídas as medidas de peso específico do ovo (g/cm3) e de peso (g) e espessura de casca (mm) e, nas análises químicas, foram avaliadas as concentrações de Ca e cinzas na casca dos ovos (%).
A análise do osso foi realizada na tíbia para avaliação de seus teores de Ca (% e g) e de cinzas (% e g).
Para o controle do consumo de ração, as rações de cada repetição dos tratamentos foram acondicionadas em baldes plásticos, devidamente identificados. O consumo foi calculado ao término de cada período de 28 dias, por meio da diferença entre a quantidade de ração fornecida e a de sobras.
A coleta dos ovos foi feita diariamente às 8 h e a produção média de ovos (expressa como a porcentagem sobre a média de aves do período - ave/dia) foi obtida computando-se os ovos inteiros, quebrados, trincados e os ovos anormais. Também foi calculado o número médio de ovos produzidos e classificados como comerciais, ou seja, sem defeitos ou trincas, determinando-se a porcentagem de postura de ovos comerciais durante o período experimental.
Para obtenção dos dados de peso médio dos ovos, todos os ovos íntegros produzidos em cada unidade experimental foram pesados em balança de precisão de 0,01 g nos três últimos dias de cada período. A massa de ovos foi calculada como a produção multiplicada pelo peso médio dos ovos.
A conversão alimentar foi obtida dividindo-se o consumo médio de ração por gramas de massa de ovos produzidos (conversão g/g) e o consumo médio de ração por dúzia de ovos produzidos (conversão g/dz).
O peso específico foi obtido por meio da coleta de todos os ovos íntegros de cada parcela no 23º, 24º e 25º dias de cada período. Para isso, inicialmente foram preparadas dez soluções de NaCl, com densidades de 1,050 a 1.100 g/cm3, com gradiente de 0,005 entre elas, previamente determinado com auxílio de um densímetro INCOTERM - OM-5565. Os ovos coletados e identificados por parcela em cada período foram então submetidos às soluções. Dos resultados obtidos nos três dias de análise, foi retirada uma média da densidade (g/cm3) dos ovos da parcela por período.
Para determinação do peso dos ovos e da espessura da casca, foram coletados aleatoriamente, nos três últimos dias de cada período de 28 dias, três ovos íntegros de cada parcela por dia. Os ovos foram identificados de acordo com o respectivo tratamento e a repetição e, posteriormente, foram pesados individualmente em balança com precisão de 0,01 g para a obtenção do peso dos ovos. Para cálculo do peso médio da casca dos ovos de cada unidade experimental, os ovos foram pesados e quebrados e suas cascas foram reservadas.
A espessura de casca, incluindo as membranas, foi medida depois que as cascas foram secas e pesadas, por meio da leitura de três pontos distintos na região equatorial utilizando-se um paquímetro digital, marca DIGIMESS, com precisão de 0,01 mm (0,01 - 25,00 mm). As três medidas obtidas em cada casca foram transformadas em um valor médio por parcela. As cascas foram secas em estufa a 105ºC e posteriormente moídas para análise dos teores de cinzas e de Ca.
Ao término do experimento, foram abatidas três aves por unidade experimental, por meio de deslocamento cervical, para retirada das tíbias direita e esquerda e determinação dos teores de Ca e cinzas nos ossos. Os ossos foram identificados por tratamento e repetição e, em seguida, foram desengordurados em extrator tipo "Soxlet" e mantidos em estufa a 55ºC durante 1 hora, sendo triturados imediatamente para as análises de cinzas e Ca utilizando-se a metodologia descrita por Silva (1998).
Os dados foram submetidos à análise de variância a 5% utilizando-se o Programa SAEG - Sistema para Análises Estatísticas e Genética (UFV, 1999). Os efeitos dos níveis de Ca foram estimados por meio de análise de variáveis pelos modelos de regressão linear e quadrática, conforme o melhor ajustamento obtido para cada variável, considerando o comportamento biológico das aves. Para a comparação entre períodos, aplicou-se o teste de média de Student Newman-Keuls (SNK) a 5% de probabilidade.
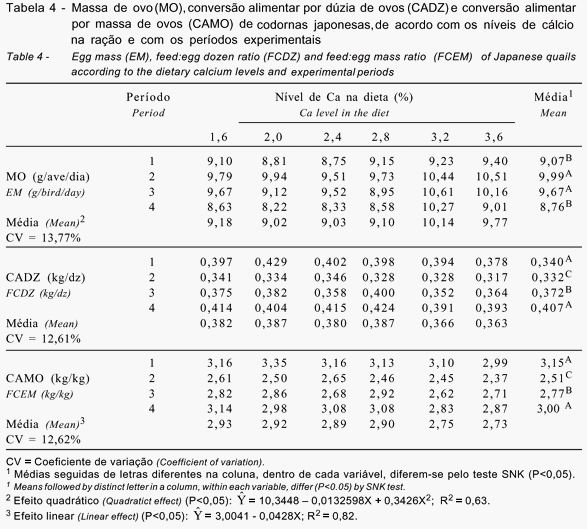
Os valores obtidos para o consumo de ração, a produção de ovos/ave/dia, a produção de ovos comercias e o peso médio do ovo são apresentados na Tabela 3 e aqueles obtidos para a massa de ovo, a conversão alimentar por dúzia de ovos e a conversão alimentar por massa de ovos são descritos na Tabela 4.
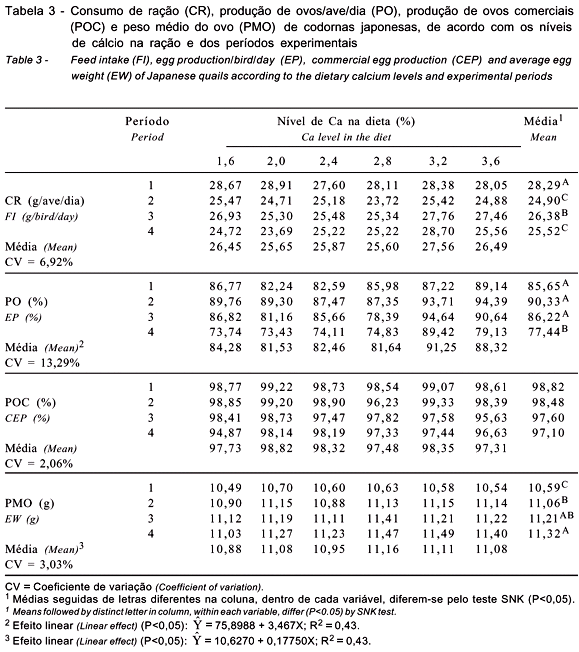
Pelas análises estatísticas, constatou-se que não houve interação significativa (P>0,05) nível de Ca da dieta × período experimental para nenhuma das variáveis analisadas, indicando que os efeitos dos níveis de Ca e dos períodos comportaram-se de modo diferente e independente.
Os resultados comprovaram que os níveis de Ca não influenciaram (P>0,05) o consumo de ração das aves (Tabela 3). Estes resultados corroboram os achados de Pedroso et al. (1999) e Masukawa et al. (1996), que testaram, respectivamente, níveis de 2,5 a 3,5% e de 2,0 a 3,5% de Ca em codornas japonesas e também verificaram que o consumo de ração não foi influenciado pelos níveis de Ca da dieta. Neste estudo, o consumo de ração foi maior no 1º período, quando as codornas formam suas reservas corporais, aumentando a produção, o peso de ovo e, conseqüentemente, a massa de ovos. O aumento no consumo de ração no 3º período pode estar relacionado à queda na temperatura ambiental durante esta fase, na qual foram registradas as menores temperaturas ao longo do dia, chegando a 13,1ºC (Tabela 2).
Verificou-se efeito linear (P<0,05) positivo da produção de ovos/ave/dia com o aumento dos níveis de Ca na dieta (Tabela 3). As dietas contendo 3,2 e 3,6% de Ca foram as que proporcionaram maior produção, no entanto, a produção obtida com o nível de 3,2% de Ca foi 3,32% superior, em valor absoluto, quando comparada àquela contendo 3,6% de Ca. Observou-se queda acentuada da produção em todos os tratamentos durante o 4º período, exceto para aqueles contendo 2,8 e 3,2% de Ca (a produção com 3,2% de Ca foi, em valor absoluto, 19,5% superior à do tratamento com 2,8% de Ca).
Avaliando quatro níveis de Ca (2,0; 2,5; 3,0 e 3,5%) e três granulometrias (fina, média e grossa) de calcário para codornas japonesas de 56 dias até 196 dias de idade, Luz (2002) observou aumento na produção de ovos com o aumento dos níveis de Ca na ração, o que está de acordo com os resultados encontrados neste experimento, em que os maiores níveis de Ca na ração proporcionaram melhor produção de ovos.
Garcia et al. (2000), trabalhando com codornas japonesas no pico de postura alimentadas com ração contendo níveis de 2,5 a 4,0% de Ca, notaram que o nível de 3,06% de Ca, próximo ao encontrado neste experimento, promoveu maximização na produção de ovos. Por sua vez, Pedroso et al. (1999) não observaram diferença significativa para porcentagem de postura, ao testarem rações contendo níveis de 2,5; 3,0 e 3,5% de Ca para codornas japonesas.
Houve efeito significativo (P<0,05) do período experimental sobre a produção de ovos/ave/dia, indicando produção menor no 4º período e maior nos três primeiros períodos, porém não diferindo significativamente entre os três primeiros. As aves ao início do 1º período experimental já estavam em elevada produção, próxima ao pico de postura, por isso, não foi observada diferença significativa entre os três primeiros períodos, o que também foi verificado por Belo (1997). A maior produção de ovos foi obtida no 2º período experimental, correspondente ao pico de postura das codornas (85 a 113 dias), seguido de decréscimo no 3º e no 4º período, comportamento esperado, tendo em vista a resposta fisiológica da ave.
Segundo Albino & Barreto (2003), codornas japonesas apresentam produção de ovos superior a 80% até aproximadamente 32ª semana de idade, quando a produção média é superior a 5,5 ovos/ave/semana. Entretanto, no último período, apenas as aves alimentadas com ração contendo 3,2% de Ca apresentaram produção superior a 80%, indicando que houve redução antecipada da produção nos demais tratamentos.
Os resultados de produção de ovos comerciais revelaram que os níveis de Ca na dieta não influenciaram (P>0,05) a produção de ovos destinados à comercialização (Tabela 3). Esses dados indicam que níveis inferiores a 2,5% de Ca na dieta, exigência determinada pelo NRC (1994), não interferiram na perda de ovos por danos na casca, pois as porcentagens de ovos viáveis à comercialização não foram significativamente diferentes entre os níveis de Ca avaliados. Nos níveis extremos (1,6 e 3,6% de Ca), a perda de ovos manteve-se praticamente a mesma, ou seja, com mesmo percentual de ovos comercializáveis.
Os níveis de Ca influenciaram significativamente (P<0,05) o tamanho dos ovos (Tabela 3). Houve aumento linear de 0,1775 g no peso do ovo para cada 1% de aumento na suplementação de Ca na dieta, conforme equação de regressão demonstrada na Tabela 3.
Houve pouca variação no peso médio dos ovos entre os tratamentos, com exceção ao menor nível de inclusão (1,6% de Ca), que resultou em menor peso médio de ovo, como esperado, pois esse nível é muito inferior ao recomendado pelo NRC (1994), que determina nível nutricional de 2,5% de Ca em dietas para codornas japonesas na fase de produção.
Em um estudo no qual testou a granulometria do calcário e os níveis de Ca descritos anteriormente, Luz (2002) não encontrou efeito significativo para peso de ovos, o que está de acordo com os resultados descritos por Pedroso et al. (1999), que utilizaram níveis de 2,5 a 3,5% de Ca em rações para codornas japonesas. Em ambos os trabalhos, foram avaliados níveis superiores a 2,0 e 2,5% de Ca, respectivamente, enquanto, neste trabalho, avaliaram-se um nível inferior aos testados por Luz (2002) e dois níveis inferiores aos avaliados por Pedroso et al. (1999). Trabalhando com níveis de 2,8 a 4,4% de Ca para poedeiras leves e semipesadas no 2º ciclo de produção, Oliveira (2001) também não encontrou efeito significativo para peso do ovo.
Todavia, Shrivastav & Panda (1986), estudando o efeito de três fontes de Ca e três níveis de Ca (3,0; 3,4; e 3,75%) para codornas japonesas de 70 até 126 dias de idade, observaram que o peso do ovo aumentou 0,4 g, em valor absoluto quando o nível de 3,0% foi aumentado para 3,75% de Ca na ração.
No decorrer do período experimental, houve aumento no peso dos ovos (P<0,05), atribuído ao desenvolvimento corporal das aves, que passaram a produzir ovos maiores e mais pesados, diminuindo a produção e aumentando o período entre as ovulações e oviposições. Verificaram-se aumentos de 4,4; 5,8 e 6,9% no peso dos ovos produzidos no 2º, 3º e 4º períodos, respectivamente, em relação aos produzidos no 1º período.
A massa diária de ovos apresentou comportamento quadrático (P<0,05) segundo a equação demonstrada na Tabela 4, indicando que maiores massas foram obtidas nos grupos de aves que receberam ração contendo 3,2 e 3,6% de Ca. Entre estes grupos, aquele alimentado com ração contendo 3,2% de Ca apresentou 3,8% a mais de massa de ovos, como resultado da maior produção de ovos/ave/dia e do maior peso de ovo, em comparação ao grupo que recebeu ração contendo 3,6% de Ca.
Estes resultados diferem dos encontrados por Shrivastav & Panda (1986), que estudaram o efeito de três fontes de Ca e de três níveis de Ca (3,0; 3,4 e 3,75%) na ração de codornas japonesas. Esses autores encontraram efeito significativo para massa de ovo, sendo que a ração contendo 3,75% de Ca resultou em redução na produção de ovos e na massa de ovo. Em pesquisa com galinhas, Keshavarz & Nakajima (1993) encontraram diferença não-significativa para massa de ovos ao fornecerem ração com níveis de 3,5 a 5,5% de Ca para linhagens de poedeiras leves e semipesadas no 2º ciclo de produção. No entanto, Abdallah et al. (1993), avaliando o desempenho de galinhas poedeiras leves e semipesadas no 2º ciclo de produção alimentadas com dietas deficientes em Ca (1,7% de Ca), abaixo da exigência (2,2% de Ca) e dieta controle (3,9% de Ca), verificaram aumento da massa de ovo com a elevação dos níveis de Ca na dieta.
Observou-se efeito (P<0,05) do período avaliado sobre a massa de ovos, sendo que maior massa foi observada no 2º e 3º períodos, que corresponderam às maiores produções e aos maiores pesos dos ovos, diferindo significativamente do 1º e do 4º períodos, que representavam a fase em que as aves já alcançaram a maturidade física e o pico máximo de produção. A redução da massa de ovo no 4º período resultou da diminuição na taxa de postura das codornas e da tendência à estabilização no tamanho dos ovos com o alcance da maturidade física das aves.
Os resultados de conversão alimentar por dúzia de ovos revelaram efeito não-significativo (P>0,05), indicando que os diferentes níveis de Ca na dieta não influenciaram este parâmetro (Tabela 4).
Estes achados corroboram os encontrados por Garcia et al. (2000), que, trabalhando com níveis de 2,5 a 4,0% de Ca em rações para codornas em postura do 70º ao 130º dia de idade, não encontraram efeito significativo para conversão alimentar por dúzia de ovos. Também está de acordo com os descritos por Masukawa et al. (2001), que verificaram efeito não-significativo para conversão alimentar por dúzia de ovos ao avaliarem níveis de 2,0 a 3,5% de Ca em rações para codornas.
Houve efeito para o período experimental (P<0,05), verificando-se melhor conversão alimentar por dúzia de ovos no 2º período, como conseqüência da maior produção de ovos e do menor consumo de ração durante o maior período de produção. A pior conversão foi para o 1º e 4º períodos, sendo que, no 1º período, as aves não haviam atingido o pico de produção, sendo observado maior consumo de ração. No 4º período, houve queda na produção de ovos e conseqüente piora na conversão alimentar das aves.
Quanto à conversão alimentar por massa de ovos, observou-se melhora de 0,043 pontos para cada aumento de 1% de Ca na dieta (P<0,05), segundo a equação indicada na Tabela 4.
As melhores conversões foram observadas nas dietas com 3,2 e 3,6% de Ca, observando-se que a dieta com 3,6% foi apenas 0,73% melhor que aquela com 3,2%. Estas melhores conversões obtidas nas dietas com 3,2 e 3,6% de Ca resultaram da maior massa de ovos, atribuída também às maiores produções e aos mais altos pesos de ovos ou à diferença não-significativa no consumo de ração entre os tratamentos.
Esses resultados estão de acordo com os descritos por Luz (2002), que encontrou melhora significativa na conversão alimentar por massa de ovos à medida que elevou os níveis de Ca na ração de codornas japonesas.
Garcia et al. (2000), no entanto, estudando níveis de 2,5 a 4,0% de Ca na dieta de codornas japonesas, não encontraram efeito para conversão alimentar por massa de ovo, resultado semelhante ao encontrado por Masukawa et al. (1996), ao trabalharem com rações contendo 2,0 a 3,5% de Ca.
As diferenças encontradas para conversão alimentar por massa de ovo (P<0,05) no decorrer dos períodos experimentais foram conseqüência das variações na produção e no peso de ovos e no consumo de ração das aves. No 1º período, a produção de ovos aumentou e as aves ainda não haviam terminado seu desenvolvimento corporal, conseqüentemente, houve maior consumo de ração, piorando a conversão alimentar neste período. Melhor conversão foi observada no 2º e 3º períodos, provavelmente porque as aves estavam no pico de produção e já haviam completado seu crescimento corporal, com produção e peso de ovos estabilizados. A partir do 4º período, a produção de ovos diminuiu e, mesmo com o maior peso dos ovos, a conversão alimentar piorou significativamente.
Os resultados referentes aos pesos médios inicial e final e ao ganho de peso das aves são apresentados na Tabela 5.
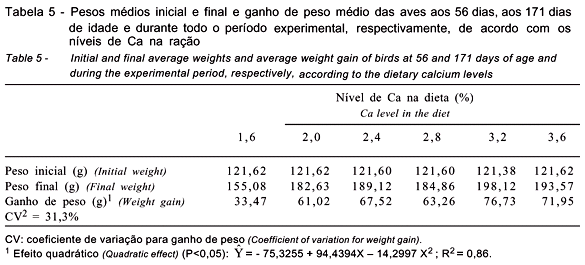
O aumento dos níveis de Ca na dieta promoveram efeito quadrático (P<0,05) sobre o ganho de peso das aves aos 171 dias de idade, indicando que maior ganho de peso foi obtido com o nível de 3,16% de Ca na dieta (Tabela 5).
Os níveis de Ca utilizados, com exceção do nível de 1,6%, proporcionaram pesos finais e ganhos de peso satisfatórios.
No último período avaliado, as reduções na produção, no peso e na massa dos ovos no tratamento com o menor nível de Ca foram de 21,3; 4,2 e 19,0%, respectivamente, em comparação aos valores obtidos com o nível de 3,2% de Ca, e de 7,3; 3,3 e 4,4%, respectivamente, em comparação aos registrados no tratamento com 3,6% de Ca. Esses resultados corroboram os encontrados por Raju et al. (1992), que verificaram aumento do peso corporal das aves com a elevação dos níveis dietéticos de Ca de 2,0 para 3,5%. Diferem, no entanto, dos obtidos por Vohra et al. (1979), que, em estudo com codornas japonesas de 147 dias a 196 dias de idade, não encontraram influência significativa para o peso corporal quando as aves foram alimentadas com ração contendo 1% de Ca. O resultado registrado por esses autores pode ser decorrente do curto período experimental, que correspondeu a apenas a 44% do período avaliado neste trabalho.
Quanto à qualidade dos ovos, não houve interação significativa níveis de Ca da ração × períodos experimentais para nenhuma das características de qualidade dos ovos: peso específico do ovo, peso da casca e espessura de casca (Tabela 6). Essa ausência de significância na interação revela que os efeitos dos níveis de Ca e dos períodos foram independentes.
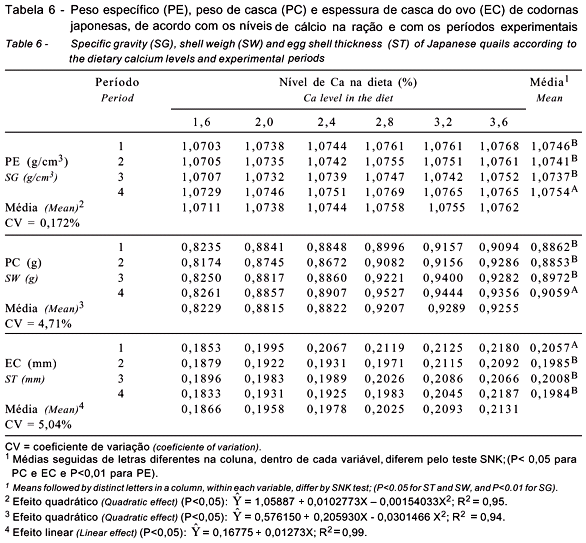
Houve efeito quadrático (P<0,01) para o peso específico dos ovos, comprovando que o melhor resultado foi observado nos ovos provenientes de codornas alimentadas com ração contendo 3,34% de Ca, segundo a equação indicada na Tabela 6. Este resultado se assemelha, em parte, ao encontrado por Luz (2002), que verificou efeito linear do peso específico de ovos de codornas alimentadas com ração contendo de 2,0 a 3,5% de Ca, indicando que o maior nível resultou em maior peso específico dos ovos. Também Pedroso et al. (1999) encontraram maior peso específico dos ovos em codornas japonesas alimentadas com dietas contendo 3,0 e 3,5% de Ca. Entretanto, Shrivastav & Panda (1986) não verificaram efeito significativo para peso específico ao testarem rações com 3,0 a 3,75% de Ca para codornas japonesas.
Nos períodos experimentais estudados, houve alteração no peso específico dos ovos com avançar da idade das aves. Os dados comprovaram não haver diferença significativa (P>0,05) entre os três primeiros períodos avaliados. Contudo, verificou-se melhora significativa (P<0,01) para esse parâmetro no último período avaliado, provavelmente em decorrência da menor produção de ovos neste período (11,3% inferior ao 3º período), resultando em melhora de 0,16% no peso específico, embora houvesse aumento significativo no peso do ovo.
Verificou-se também efeito quadrático (P<0,05) dos tratamentos sobre o peso de casca dos ovos, indicando que o maior peso foi registrado para ovos de aves alimentadas com dieta contendo 3,42% de Ca (Tabela 6). Esses resultados confirmam, parcialmente, os encontrados por Abdallah et al. (1993), que, trabalhando com poedeiras comerciais leves e semipesadas alimentadas com rações contendo diferentes níveis de Ca, observaram aumento linear do peso de casca dos ovos de ambas as linhagens, indicando redução do peso da casca nos ovos provenientes das poedeiras alimentadas com a ração com nível insuficiente de Ca.
Entre os períodos experimentais, observou-se aumento do peso da casca a partir do 3º período, que não diferiu estatisticamente do 2º período, provavelmente em razão das reduções na produção e na massa de ovos e da estabilidade de peso dos ovos com o avançar da idade das aves, que se encontravam no primeiro terço do ciclo de produção.
Os resultados comprovaram que a espessura de casca dos ovos foi influenciada linearmente (P<0,01) com o aumento dos níveis de Ca na dieta (Tabela 6). A equação indicada na Tabela 6 indica que, a cada aumento de 1% de Ca na dieta, ocorreu melhora de 0,013 mm na espessura da casca. Para a indústria alimentícia, que processa os ovos principalmente como conserva (agrega valor ao produto), os ovos com espessura de casca intermediária são mais viáveis porque apresentam maior facilidade e rapidez na remoção da casca, sem que haja rompimento da membrana interna, resultando em ovos com albúmem mais íntegros e de melhor aparência depois de descascados. Portanto, pode-se considerar viável a espessura da casca dos ovos de codornas que consumiram a ração contendo 3,2% de Ca, visto que a diferença na espessura de casca entre os ovos destas aves foi pequena em comparação aos daquelas alimentadas com ração contendo 3,6% de Ca, que foram apenas 1,1% superiores.
Analisando o efeito de fontes e níveis de Ca para codornas japonesas, Shrivastav & Panda (1986) encontraram efeito quadrático para a espessura de casca com o aumento dos níveis de Ca na dieta e determinaram exigência nutricional de 3,0% de Ca para obtenção de melhor espessura de casca do ovo. Os resultados divergem, no entanto, dos relatados por Pedroso et al. (1999), que não encontraram efeito significativo para espessura de casca de ovos de codornas recebendo dieta com 2,5 a 3,5% de Ca.
A espessura de casca diferiu (P<0,05) com o período experimental, indicando que os ovos produzidos no 1º período apresentaram maior espessura de casca quando comparados aos dos demais períodos. Entretanto, não foi observada diferença significativa entre os ovos produzidos nos três últimos períodos.
Como demonstrado na Tabela 7, os níveis de Ca na dieta não influenciaram (P>0,05) os teores cinzas e de Ca na casca dos ovos mesmo com a utilização de dietas com níveis de Ca abaixo do recomendado pelo NRC (1994), como aquelas contendo 1,6; 2,0 e 2,4% de Ca. Abdallah et al. (1993), no entanto, trabalhando com poedeiras alimentadas com diferentes níveis de Ca (1,7 a 3,9%), verificaram maior deposição destes minerais na casca dos ovos quando aumentaram os níveis de Ca na dieta das aves.
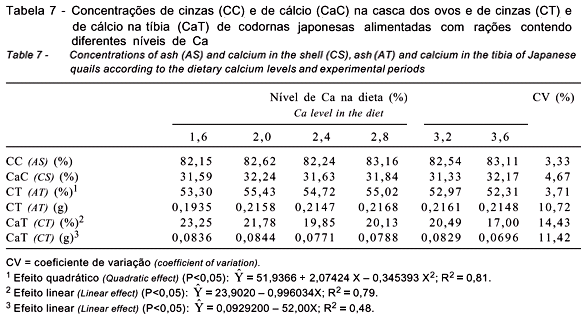
À exceção do teor de cinzas no osso em gramas, as concentrações de cinzas (em porcentagem) e de Ca (em gramas e em porcentagem) nos ossos foram afetadas (P<0,05) pelos níveis de Ca estudados (Tabela 7). Houve comportamento quadrático (P<0,05) para o teor de cinzas, em porcentagem, nos ossos, sendo que a maior concentração foi obtida com o nível de 3,0% de Ca na ração. Os resultados diferiram daqueles apresentados por Luz (2002), que não encontrou efeito significativo para porcentagem de cinzas nos ossos de codornas ao avaliar níveis de 2,0 a 3,5% de Ca na ração e também são diferentes dos observados por Garcia et al. (1999), que não verificaram efeito significativo para porcentagem de cinzas nos ossos quando testaram níveis de 2,5 a 4,0% de Ca na ração.
Verificou-se redução linear (P<0,05) da concentração de Ca no osso, em porcentagem e em valor absoluto, com o aumento do nível de Ca da ração (Tabela 7). Esses resultados diferem dos achados por Garcia et al. (2000), que não notaram efeito significativo para a porcentagem de Ca nos ossos de codornas com o aumento dos níveis de Ca na ração. Também são diferentes dos reportados por Abdallah et al. (1993), que verificaram aumento desta concentração quando elevaram o nível de Ca da dieta de 1,7 a 3,9% de Ca para poedeiras leves e semipesadas na fase de produção. O comportamento da concentração de Ca na tíbia observado neste trabalho foi inesperado, pois o menor consumo diário de Ca resultou em maior sedimentação, em vez de maior reabsorção, deste mineral nos ossos.
De modo geral, os níveis de 3,2 e 3,6% de Ca proporcionaram melhor desempenho produtivo e melhor qualidade dos ovos das codornas. Estes níveis são superiores ao apresentado pelo NRC (1994), que cita o nível de 2,5% de Ca na dieta como o mais adequado para codornas japonesas em postura. No entanto, sugere-se o fornecimento de 3,2% de Ca, pois a ração de codorna é mais energética e mais protéica que a de poedeiras comerciais e, neste caso, uma dieta contendo 3,6% de Ca necessitaria de maior quantidade de calcário como fonte de Ca e de maior quantidade de óleo como fonte de energia, o que resultaria em maior custo de produção.
A exigência de Ca estimada para codornas japonesas na fase inicial de produção (56 a 171 dias de idade) variou de 3,0 a 3,6% na dieta para o atendimento aos parâmetros avaliados. Portanto, respeitando-se o comportamento biológico das aves, recomenda-se o nível de 3,2% de Ca na ração, que corresponde a um consumo diário de 882 mg de Ca/ave, ou de 87 mg de Ca/g de ovo, como exigência para obtenção de maior produção, melhor conversão alimentar (massa e dúzia de ovos) e manutenção da qualidade dos ovos.
ABDALLAH, G.; HARMS, R.H.; EL-HUSSEIN, O. Performance of laying eggs with heavy or light shell weight when fed diets with different calcium and phosphorus levels. Poultry Science, v.72, p.1881-1891, 1993.
ALBINO, L.F.T.; BARRETO, S.L.T. Criação de codornas para produção de ovos e carne. Viçosa, MG: Aprenda Fácil, 2003. 289p.
BAILEY, C.A.; LINTON, S.; BRISTER, R. et al. Effects of grade levels of dietary phosphorus on bone mineralization in the very young poultry. Poultry Science, v.65, p.1018-1020, 1986.
BELO, M.T.S. Níveis de energia metabolizável e de metionina em rações de codornas japonesas (Coturnix coturnix japonica) na fase inicial de produção. Lavras: Universidade Federal de Lavras, 1997. 60p. Tese (Mestrado em Zootecnia) - Universidade Federal de Lavras, 1997.
BRESSAN, M.C.; ROSA, F. C. Processamento e industrialização de ovos de codornas. In: SIMPÓSIO INTERNACIONAL DE COTURNICULTURA - Novos CONCEITOS APLICADOS À PRODUÇÃO DE CODORNAS, 1., 2002, Lavras. 2002. Anais... Lavras: Universidade Federal de Lavras, 2002. p.68-69.
FROST, T.J.; ROLAND, D.A. The influence of various calcium and phosphorus levels on tibia strength and eggshell quality of pullets during peak production. Poultry Science, v.70, n.4, p.963-969, 1991.
FURLAN, A.C.; ANDREOTTI, M.O.; MURAKAMI, A.E. et. al. Valores energéticos de alguns alimentos determinados com codornas japonesas (Coturnix coturnix japonica) In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1996, Curitiba. Anais... Campinas: Fundação Apinco de Ciência e Tecnologia Avícolas, 1996. p.43.
GARCIA, J; MURAKAMI, A.E.; MARTINS, E.N. et al. Exigências nutricionais de cálcio e fósforo para codornas japonesas (Coturnix coturnix japonica) em postura. Acta Scientiarum, v.22, n.3, p.733-739, 2000.
HAMILTON, R.M.G.; CIPERA, J.D. Effects dietary calcium levels during the brooding, rearing, and early laying period on feed intake, egg production, and shell quality of white leghorn hens. Poultry Science, v.60, p.349-357, 1981.
INSTITUTO NACIONAL DE LA RECHERCHÉ AGRONOMIQUE - INRA. Alimentação dos animais monogástricos: suínos, coelhos e aves. 2.ed. São Paulo: Roca, 1999. 245p.
KESHAVARZ, K.; NAKAJIMA, S. Re-evaluation of phosphorus requirements of laying hens for optimum performance and eggshell quality. Poultry Science, v.72, p.114-153, 1993.
LEESON, S.; JULIAN, R.J.; SUMMERS, J.D. Influence of prelay and early-lay dietary calcium concentration on performance and bone integrity of leghorn pullets. Canadian Journal Animal Science, v.66, p.1087-1095, 1986.
LUZ, L.C.P. Variação granulometrica do calcário e diferentes níveis de cálcio em rações de codornas japonesas (Coturnix coturnix japonica). Lavras: Universidade Federal de Lavras, 2002. 58p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Lavras, 2002.
MASUKAWA, Y.; MORAES, V.M.B.; ARIKI, J. et al. Efeito dos níveis de cálcio sobre o desempenho produtivo e qualidade dos ovos de codornas japonesas (Coturnix coturnix japonica) In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1996, Curitiba. Anais... Curitiba: Fundação Apinco de Ciência e Tecnologia Avícolas, 1996. p.35.
MASUKAWA, Y.; MORAES, V.; M.B.; ARIKI, J. et al. Níveis de cálcio da dieta sobre o desempenho e a qualidade da casca de ovos de codornas japonesas. Ars Veterinária, v.17, n.2, p.144-148, 2001.
NATIONAL RESERCH COUNCIL - NRC. Nutrient requirements of poultry. 9.ed. Washington, D.C.: National Academy of Sciences: 1994. 155p.
OLIVEIRA, J.R. Níveis de cálcio e fósforo em dietas para poedeiras leves e semipesadas no 2º ciclo de produção. Lavras: Universidade Federal de Lavras, 2001. 61p. Dissertação (Mestrado em Zootecnia) - Universidade Federal de Lavras, 2001.
PEDROSO, A.; MORAES, V.M.B.; ARIKI, J. et al. Efeito de níveis dietéticos de cálcio e fósforo disponível sobre o desempenho e qualidade dos ovos de codornas japonesas. Ars Veterinária, v.15, n.2, p.135-139, 1999.
PINTO, R.; DONZELE, J.L.; FERREIRA, A.S. et al. Exigência de metionina mais cistina para codornas japonesas em postura. Revista Brasileira de Zootecnia, v.32, n.5, p.1166-1173, 2003a.
PINTO, R.; FERREIRA, A.S.; DONZELE, J.L et al. Exigência de lisina para codornas japonesas em postura. Revista Brasileira de Zootecnia, v.32, n.5, p.1182-1189, 2003b.
RAJU, M.V.L.N.; RAO, P.V.; REDDY, V.R. Effect of dietary calcium and inorganic phosphorus on the performance of laying Coturnix quail. Indian Journal of Animal Science, v.62, p.1072-1076, 1992.
ROSTAGNO, H.S.; ALBINO, L.F.T.; DONZELE, J.L. et al. Composição de alimentos e exigências nutricionais de aves e suínos (Tabelas brasileiras). Viçosa, MG: Universidade Federal de Viçosa, 2000. 141p.
SHRIVASTAV, A.K. Recentes avanços na nutrição de codornas japonesas. In: SIMPÓSIO INTERNACIONAL DE COTURNICULTURA - NOVOS CONCEITOS APLICADOS À PRODUÇÃO DE CODORNAS, 1., 2002, Lavras. Anais... Lavras, 2002, p.116-117.
SHRIVASTAV, A.K.; PANDA, B. Level an sources of calcium for egg production and Shell quality in quails. Indian Journal Poultry Science, v.21, n.1, p.78-81, 1986.
SHRIVASTAV, A.K.; PANDA, B.; DARSHAN, N. Calcium and phosphorus requirements of laying Japanese quails. Indian Journal Science, v.24, n.1, p.27-34, 1989.
SILVA, D.J. Análises de alimentos (Métodos químicos e biológicos). Viçosa, MG: Universidade Federal de Viçosa, 1998. 166p.
STADELMAN, J.W.; COTTERILL, O.J. Egg science and technology. 4.ed. New York: Food Products Press, 1995. 590p.
UNIVERSIDADE FEDERAL DE VIÇOSA - UFV. Manual de utilização do programa SAEG (Sistema para Análises Estatísticas e Genéticas). Viçosa, MG: Central de Processamento de Dados, 1999. 59p.
VOHRA, P.; SIOPES, T.D.; WILSON, W.O. Egg production and body weight changes in japanese quail and leghorn hens following deprivation of either supplementary calcium or vitamin D. World's Poultry Science Journal, v.48, n.1, p.5-16, 1979.
Sergio Luiz de Toledo BarretoI;
sbarreto@ufv.br
Ciane Aparecida PereiraII;
Regina Tie UmigiII;
Tatiana Cristina da RochaII;
Marcelle Santana de AraujoII,
Cynthia Siqueira SilvaIII;
Rodolpho de Almeida Torres FilhoIV
IUniversidade Federal de Viçosa - Departamento de Zootecnia, s/n - Campus Universitário - Viçosa - MG
IIMestrando em Zootecnia, Universidade Federal de Viçosa
IIIMestrando em Zootecnia, Universidade Estadual Fluminense - RJ
IVDoutorando em Zootecnia, Universidade Federal de Viçosa MG
Revista Brasileira de Zootecnia v.36 n.1 Viçosa jan./fev. 2007
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




