La gametogénesis ocurre en las glándulas
sexuales o gónadas, tanto en machos como
hembras cuando ellos han alcanzado la madures sexual. Este
momento conocido como pubertad, se caracteriza en los
mamíferos por un desencadenamiento de una
liberación basal continua de gonadotrofinas (FSH y LH) en
los machos, o una liberación cíclica de ellas en
las hembras. Estas hormonas actúan sobre sus
órganos blancos, las gónadas, activando tanto la
gametogénesis como la síntesis de esteroides
gonadales. Estos esteroides actúan a su vez sobre las
características sexuales tanto secundarias como
accesorias, provocando los cambios típicos observados
durante la pubertad.
En términos de gametogénesis, la pubertad
tiene un significado totalmente diferente en el macho y en la
hembra. En el macho ella coincide con la división
redaccional y con las primeras ondas de
espermiación.
En la hembra, por acción del balance
gonadotrofico presente en esta etapa, se reanuda la meiosis, se
completa la división redaccional y ocurre la
ovulación, la cual tiene lugar en periodos bien definidos
de cada ciclo sexual.
Entre los procesos de espermatogénesis y
ovogénesis existe una diferencia esencial: la
ovogénesis no es un proceso continuo como lo es la
espermatogenesis. Las ovogonias entran en profase meiótica
en una edad muy temprana la cual se detiene en un estado llamado
dictioteno o de reposo. La célula resultante, el ovocito
I, rodeado de las células somáticas
(pregranulosicas), pasa a llamarse folículo primordial. El
conjunto de estos folículos forman la llamada
"población de folículos de reserva" a partir de la
cual, en cada ciclo, un numero determinado de ellos se moviliza y
crece alcanzando el estado de folículo preovulatorio o
sigue el camino de la atresia (degeneración).
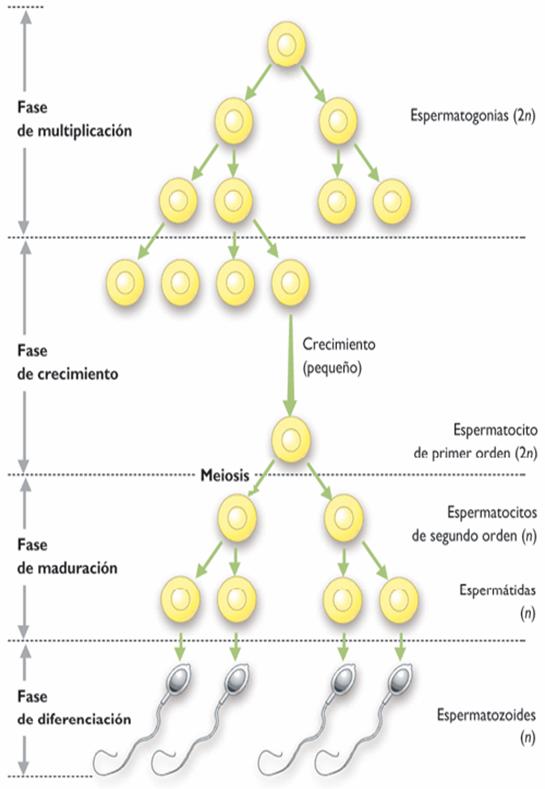
La gametogénesis comprende 4 fases
1. Origen y migración de las
gónadas.
2. Multiplicación de las
células germinales
3. Meiosis
4. Maduración final antes de la
fertilización
3.
ESPERMATOGÉNESIS
La espermatogénesis es la suma de los procesos
que en el adulto terminan con la producción de
espermatozoides, siendo por ende un continuo que se inicia ya en
la vida embrionaria. Se produce por la activación del eje
hipotalámico-hipofisiario que involucra la
secreción de la hormona estimulante del folículo y
la hormona luteinizante.
Existe espermatogénesis longitudinal en insectos
y peces, espermatogenesis quística en anfibios urodelos y
espermatogénesis, radial en reptiles y
mamíferos.
La formación de los espermatozoides comienza en
la pubertad y ocurre en el interior de los tubos
seminíferos mediante tres etapas.
3.1. Prenatal o período proliferativo:
En la tercera semana de gestación se pueden
reconocer las células primordiales germinales,
éstas se dirigen hacia el lugar donde están los
testículos y allí sufren una división
mitótica que da origen a los gonocitos, estos se
multiplican y se originan las espermatogonias, que se ubican en
la base del tubo seminífero.
3.2. Posnatal o período de
crecimiento:
Después del nacimiento, las espermatogonias
fetales dan origen a las espermatogonias prepuberales o
espermatocitos primarios, los cuales son células diploides
(2N), éstas son las que originan los espermatozoides que
se forman al pasar la pubertad.

3.3. Madurez Sexual o período de
maduración: Cuando se activa el sistema
hipotalámico-hipofisiario se produce la
formación de los espermatozoides; las espermatogonias
mediante la mitosis forman los espermatocitos de primer orden,
éstos pasan por un proceso de meiosis, antes de la primera
división meiótica los espermatocitos de primer
orden aumentan de tamaño y sus cromosomas se duplican. Las
células resultantes de la primera división
meiótica se llaman espermatocitos de segundo orden, luego
ocurre la segunda división meiótica y se originan
las espermatidas, éstas sufren modificaciones y producen
los espermatozoides.
4.
ANATOMÍA E HISTOLOGÍA DEL
TESTICULO
El aparato reproductor masculino consiste en la
gónada o testículo y en los órganos sexuales
accesorios. Entre estos se encuentran los conductos de
eliminación de semen, una serie de glándulas anexas
que contribuyen a la secreción de la parte liquida del
semen y el pene.
Los testículos son órganos ovoides pares,
situados en el escroto. El testículo esta rodeado por una
firme envoltura fibrosa, la albugínea. En su cara interna
o tunica vasculosa es menso densa y contiene numerosos vasos. En
la región posterocefalica del testículo, esta
tunica forma el mediastino testicular por le cual entran al
órgano los conductos, vasos y nervios. Desde allí
irradian tabiques de tejido conectivo que dividen al
parénquima testicular en alrededor de 250
lóbulos.
La arteria testicular es extremadamente flexuosa y da
lugar a una irrigación de tipo terminal. Los plexos
venosos y en especial los plexos pampiniformes merecen especial
mención como mecanismo termorregulador de la sangre
intratesticular. Los espacios linfáticos son muy amplios y
en muchas especies constituyen gran parte del área
intertubular. Los nervios de tipo vasomotor y sensitivo inervan
los vasos y el tejido intersticial o le Leydig. Cabe destacar que
en los mamíferos llegan solo a cierta distancia de los
túbulos, sin penetrar en el compartimiento
peritubular.
Cada lóbulo testicular contiene de uno a tres
túbulos muy flexuosos, los túbulos
seminíferos. Estos tiene la forma de un arco, con ambos
extremos conectados, en la zona del mediastino testicular, ala
rete testis. Esta constituye el inicio del sistema excretor de
gametos y fluido testicular.
Cada túbulo mide alrededor de 80 cm de largo y
unos 200 a 250 um de diámetro. El largo total de los
túbulos seminíferos en el hombre es de unos 250
metros. El túbulo esta rodeado de un tejido denominado
peritubular, y en su interior contiene celular somáticas o
de Sertoli y progenie de células germinales organizada en
el epitelio seminífero.
En suma, en la organización testicular podemos
distinguir los siguientes compartimentos: a) compartimento
intratubular, formado por células somáticas (de
Sertoli) y germinales (que constituyen el epitelio
seminífero); b) compartimento peritubular,
con células especializadas (contráctiles o mioides)
y sus productos intercelulares; c) compartimento
extratubular, que comprende los subcompartimentos vascular
(arterial venoso), linfático, intersticial o conectivo
propiamente tal e intersticial endocrino (células de
Leydig).
5. CITOLOGIA DE
LA ESPERMATOGÉNESIS
5.1. ESPERMATOGONIAS
Las espermatogonias se pueden localizar junto a las
células de Sertoli, cercanas a la membrana basal, estas
son las únicas células germinativas que se
presentan antes de la pubertad., de un testículo infantil,
se puede apreciar en la pared de los túbulos
seminíferos dos tipos celulares, las células de
Sertoli y las espermatogonias.
Las espermatogonias se dividen en dos variedades: de
tipo A y tipo B
Las espermatogonias tipo A son menos numerosas y de
forma aproximadamente oval, parcialmente rodeada por la
célula de Sertoli. El núcleo es oval y el
nucleoplasma es pálido, presentando una fina
granulación. El citoplasma es pobre en organitos
membranosos. Su división es por mitosis y casi la mitad de
las células hijas permanecen como tipo A que actúan
como células madres, mientras que el resto se transforma
en espermatogonias de tipo B.
El tipo B de espermatogonia es mucho más numeroso
y se localiza también parcialmente rodeado por las
células de Ser-toli, o en contacto con otras
espermatogonias B. El cuerpo celular es redondeado, con un
núcleo esférico en el que se observan grumos de
cromatina y un nucleolo heterogéneo e irregu-lar. El
citoplasma es comparable con el de la espermatogonia A y en
él también se observan partículas que
parecen corresponder-se con polirribosomas.
Al final de la última división de estas
células, la división del cuerpo celular es
incompleta, persistiendo puentes intercelulares en las siguientes
divisiones meióticas, manteniendo unidos hasta cuatro
espermatocitos secundarios y ocho espermátidas.
Al final de la última división de las
espermatogonias B, las células hijas son los
espermatocitos primarios. Estas células ocupan
aproximadamente la zona media del epitelio, y es en ellas que
ocurre la división meiótica.
Las características morfológicas de dichas
células están en relación con el
estadio fisiológico en que se encuentren. En
el citoplasma se puede observar un mayor desarrollo de los
organitos membranoso. Una característica importante es la
presencia de puentes intercelulares.
5.2. ESPERMATOCITOS
Los espermatocitos primarios se hallan en estrecho
contacto con las células de Sertoli. Producto de la
primera división de maduración de la meiosis, que
ocurre en el espermatocito primario, surgen los espermatocitos
secundarios, en los cuales continúa el proceso de
división. Estas son células pequeñas que se
dividen con rapidez, para dar lugar a las células con un
número haploide de cromosomas que son las
espermátidas.

5.3. ESPERMÁTIDAS
Las espermátidas son células
pequeñas, aproximadamente esféricas, y de
núcleo también esférico. Muestran gran
desarrollo de algunos organitos membranosos, tales como
mitocondrias y aparato de Golgi, pero también la
morfología del citoplasma va a estar en relación
con el estado de espermiogénesis en que se encuentra la
célula
6.
ESPERMIOGÉNESIS
En la espermiogénesis, los principales cambios
que ocurren en las espermátidas son los
siguientes:
1. El aparato de Golgi, situado muy cercano al
núcleo, comienza a formar vesículas que se unen
formando una vesícula acrosómica que se adosa a la
membrana externa de la envoltura nuclear. Esta vesícula
sigue creciendo por fusión de otras que provienen del
aparato de Golgi, y van rodeando la parte superior del
núcleo, hasta cubrir la mitad de él; posteriormente
se condensa el material acrosómico y la membrana de la
vesícula forma en el núcleo el capuchón
cefálico.
2. De forma simultánea a lo anterior, en un polo
del núcleo (polo opuesto) comienza a desarrollarse el
flagelo, órgano que sirve para el desplazamiento del
espermatozoide En estadios tempranos del desarrollo del flagelo
aparece el manchete, que es una cortina de microtúbulos
que rodea la parte inferior del núcleo y participa en los
cambios de formas nucleares y en la implantación del
flagelo.
3. La masa excesiva de citoplasma que va quedando
según se forma el espermatozoide, contiene muchos
mitocondrias y estas rodean la pieza media del flagelo formando
la vaina mitocondrial, la cual proporciona la energía para
su movimiento.
7.
ESPERMATOZOIDE
Los espermatozoides están compuestos
por:
La cabeza, que contiene al pronúcleo portador de
la información genética, el acrosoma contenedor de
enzimas hidrolíticas que degradan la zona pelucida, una
estructura glicoproteica que rodea al ovocito, y una delgada capa
de citoplasma, todo ello envuelto en una membrana
plasmática que lo une al cuello. Tiene una media de 4 o 5
&µm de largo.
El cuello es corto y estrecho. Contiene una placa basal
que lo separa de la cabeza y los centríolos modificados.
De uno de ellos (el distal) se origina el flagelo.
La pieza media (de unos 4 o 5 &µm de
longitud), posee una gran cantidad de mitocondrias concentradas
en una vaina helicoidal, que proveen de energía al
espermatozoide producciendo ATP.
La cola, que le proporciona movilidad (zona
flagélica funcional recubierta sólo de
membrana

8. CÉLULAS
SUSTENTACULARES O DE SERTOLI
Las células de Sertoli ocupan la mayor parte del
espesor de la pared de los túbulos, encontrándose
las bases de estas células en contacto directo con la
membrana basal.
Por su forma irregular no es posible observar con
claridad los límites de estas células al M/O. Los
contornos celulares son imprecisos, ya que las células
germinales, durante su diferenciación, ocupan profundas
depresiones en el citoplasma de las células de Sertoli,
las cuales no solo sirven de sostén, sino que
también participan en su nutrición.
La célula de Sertoli es una célula
columnar alta, que presenta su núcleo cerca de la
región basal, mientras que el resto del citoplasma se
extiende hasta llegar a veces cerca de la luz del
túbulo.
El núcleo observado al M/E es generalmente
ovoide, pero presenta pliegues profundos en su contorno. El
nucleoplasma es relativamente homogéneo, y lo más
característico que presenta es el nucleolo.
El citoplasma presenta un aspecto reticular que contiene
delgados filamentos y microtúbulos aislados o en
ordenamiento paralelo al eje mayor de la
célula.
Las mitocondrias son alargadas y bastante numerosas y el
REL está mucho más desarrollado que el RER. Se
observan también gotas de lípidos y delgados
cristales fusiformes, localizados cerca del núcleo. El
significado fisiológico de estos cristales se desconoce,
aunque son característicos en el humano.
9. CÉLULAS
INTERSTICIALES Ó DE LEYDIG.
Como planteamos al principio del capítulo, las
células intersticiales ó de Leydig se localizan en
el tejido intersticial y pertenecen al parénquima, pues
tienen la importante función de comportarse como
glándulas endocrinas puesto que secretan las hormonas
sexuales masculinas (testosterona).
Estas células epitelioides se encuentran
agrupadas formando acúmulos en estrecha relación
con los vasos sanguíneos. Las células de Leydig son
células grandes de forma poliédrica irregular, de
núcleo excéntrico que contiene gránulos de
cromatina y un nucleolo bien desarrollado. El citoplasma, de
aspecto vacuolado, presenta abundantes inclusiones en forma de
gotas lipídicas; en el humano presentan los denominados
cristaloides de Reinke cuyo significado fisiológico se
desconoce.
El organito más desarrollado es el REL, lugar de
síntesis de las hormonas esteroides; las mitocondrias
muestran crestas tubulares características de las
células que producen estas hormonas.
10. SISTEMA DE
CONDUCTOS EXCRETORES
Los conductos excretores del sistema reproductor
masculino están constituidos por estructuras como los
tubos rectos (que parten de los túbulos
seminíferos), la red testicular o rete testis, los
conductillos eferentes, el epidídimo y los conductos
deferentes y eyaculadores.
10.1. Tubos rectos. Son cortos y parten del
vértice de cada lobulillo testicular; están
formados por un epitelio cúbico simple, en el que
sólo se observan células de Sertoli, separadas del
tejido conjuntivo laxo por una lámina basal.
10.2. Rete testis o red testicular. Los tubos rectos
llegan al mediastino testicular donde forman parte de una red de
tubos anastomosados formados por un epitelio cúbico
simple.
10.3. Conductillos eferentes. De la red testicular
parten de 12 a 15 conductillos eferentes formados por un epitelio
cilíndrico simple ciliado. Por debajo de la membrana basal
del epitelio se observa una delgada capa de músculo liso
en disposición circular. Estos conductillos se hacen muy
espiralados y constituyen los lobulillos del
epidídimo.
10.4. Epidídimo. Cada epidídimo tiene una
cabeza, un cuerpo y una cola, y presenta una cubierta fibrosa
similar a la albugínea. La cabeza, en su mayor parte,
está constituída por los conductillos eferentes
unidos mediante un tejido conjuntivo muy
vascularizado.
Los conductillos eferentes desembocan posteriormente en
el conducto del epidídimo, formado por un revestimiento
epitelial, una membrana basal y una delgada capa de fibras
musculares lisas, dispuestas circularmente. El epitelio a todo lo
largo del conducto es cilíndrico seudoestratificado con
dos tipos de células, las pequeñas basales y las
cilíndricas altas que presentan estereocitos. El conducto
del epidídimo comprende la mayor parte del cuerpo y de la
cola, y en su porción distal, se va haciendo cada vez
más recto hasta que se continúa con el conducto
deferente.
10.5. Conducto deferente. La pared del conducto
deferente es gruesa y su luz es estrecha, pues la mucosa forma
pliegues longitudinales que determinan el contorno irregular de
la luz. El epitelio que lo reviste es seudoestratificado y
presenta células altas que poseen estereocilios; por
debajo se encuentra la membrana basal que lo separa de la
lámina propia. Por debajo de la mucosa se localiza la
submucosa con muchos vasos sanguíneos e inmediatamente se
presenta la capa muscular, que es la más gruesa y presenta
las fibras dispuestas en tres direcciones. La más cercana
a la submucosa se orienta longitudinalmente, la capa media es
circular y la más externa es nuevamente
longitudinal; esta última se encuentra directamente
en contacto con la adventicia que se une a los
tejidos vecinos.
La porción final dilatada del conducto presenta
una luz más amplia y mayor cantidad de pliegues en la
mucosa y constituye la ampolla del conducto deferente.
Este conducto, junto con la arteria espermática,
el plexo venoso pampiniforme y los nervios del plexo
espermático y fibras longitudinales de músculo
estriado (músculo cremaster) forman el cordón o
fascículo espermático.
10.6. Conductos eyaculadores. Estos conductos se forman
por la unión de la extremidad distal de la ampolla del
conducto deferente y el conducto excretorio de la vesícula
seminal. La mucosa forma pliegues delgados que se proyectan hacia
la luz del conducto. El epitelio de la mucosa es
cilíndrico simple ó seudoestratificado.
11.
Glándulas anexas
Las glándulas anexas del sistema reproductor
masculino, son la próstata, las vesículas seminales
y las glándulas bulbouretrales.
11.1. Próstata
La próstata rodea la uretra, cuando esta sale de
la vejiga. Es una glándula tubuloacinar de forma
cónica y aplanada en sentido anteroposterior, siendo
más gruesa en la parte superior. Tiene consistencia firme
y está rodeada por una cápsula delgada de tejido
conjuntivo, de la cual parten tabiques anchos y gruesos que
separan ampliamente las glándulas, presenta además
fibras musculares lisas.
El tejido parenquimatoso está representado por un
conglomerado de 30 a 50 glándulas pequeñas, que se
hallan distribuidas en tres zonas bien delimitas, situadas
más o menos concéntricas y rodeando a la
uretra.
La región prostática que está por
delante de la uretra está prácticamente desprovista
de glándulas y la parte situada entre la uretra y los
conductos eyaculadores contiene glándulas que se abren
cerca de la abertura utrículo prostático. Este es
un divertículo ciego glandular que se extiende hasta el
lóbulo medio de la próstata entre los conductos
eyaculadores.
En la zona media se localizan las glándulas
submucosas, y por último, en la parte más
externa que ocupa mayor espacio en la glándula, se
localizan las glándulas principales que producen la mayor
parte de la secreción prostática.
El epitelio glandular es de tipo cilíndrico
simple o seudoestratificado y descansa sobre una capa de tejido
conjuntivo con muchas fibras elásticas y células
musculares lisas.
Las unidades secretoras, en sus extremos ciegos, son de
menor calibre que el resto de la unidad. Por lo cual en los
cortes se evidencian porciones amplias que alternan con tubos
estrechos ramificados. Las células del epitelio secretor
presentan abundantes mitocondrias y un aparato de Golgi
desarrollado.
La secreción prostática es un
líquido poco viscoso con bajo contenido proteíco,
pero es la fuente principal del ácido cítrico y de
la fosfatasa ácida del semen. En la luz de las
glándulas se observan frecuentemente acúmulos de
material secretor, denominados concreciones
prostáticas.
11.2. Vesículas seminales
Estos órganos están localizados
simétricamente a cada lado de la línea media, por
detrás de la vejiga y por encima de la próstata.
Son tubos largos muy apelotonados y enrollados sobre sí,
cuya pared presenta tres capas:
1. Capa que rodea la luz del túbulo. Es la mucosa
que presenta numerosos pliegues, lo que amplía la
superficie secretora. El epitelio varía con la edad y las
condiciones fisiológicas, pero es generalmente
seudoestratificado formado por una capa de células basales
y otra capa de células cilíndricas. La
lámina propia parece contener glándulas, pero eso
se debe solamente a los pliegues de la mucosa.
2. Capa media. Es muscular, con las fibras lisas
dispuestas circularmente en la parte interna, y longitudinales en
la parte externa.
3. Capa externa. Es la más externa y está
formada por tejido conjuntivo con muchas redes de fibras
elásticas. Las vesículas producen un líquido
viscoso de color amarillo.
La estructura del epitelio de las vesículas
seminales varía según las condiciones
hormonales, por lo que se modifica con la edad del
individuo.
11.3. Glándulas bulbouretrales
Las glándulas bulbouretrales están
situadas en el compartimiento perineal profundo, entre el margen
inferior de la próstata y el bulbo de la uretra. Las
glándulas bulbo uretrales de Cowper son túbulos
alveolares compuestas y su secreción es de tipo mucoso.
Los conductos y las unidades secretoras son de forma y
tamaño irregular y la estructura del epitelio varía
según el estado funcional, encontrándose desde
células aplanadas en los alvéolos dilatados hasta
células cúbicas y cilíndricas.
El epitelio de los conductos es de tipo
seudoestratificado, similar al de la uretra. La secreción
es viscosa y clara, muy parecida a la mucina.
A su salida de los tubos seminíferos, los
espermatozoides no tienen aún capacidad fecundante ni
motilidad, sólo viajan por las primeras porciones del
sistema de conductos, ayudados por las contracciones de las
paredes de estos tubos, debido a la actividad del músculo
liso. Es a nivel del epidídimo que adquieren las dos
propiedades antes mencionadas.
Durante la eyaculación, al paso de los conductos
se une el líquido seminal a los espermatozoides. Dicho
líquido es producido por las glándulas anexas al
aparato reproductor y junto con los espermatozoides forma el
semen que es expulsado durante la eyaculación mediante el
órgano copulador, el pene.
12.
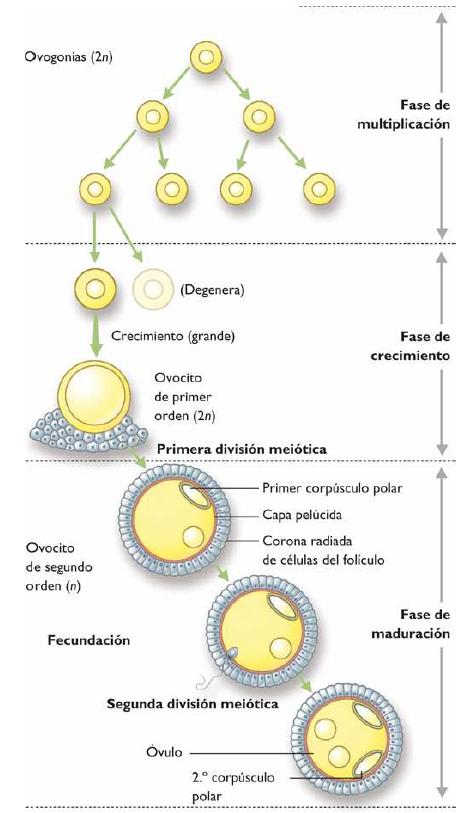
OVOGÉNESIS:
Este proceso se inicia en la mayoría de las
hembras de mamíferos durante la etapa embrio-fetal. Solo
en las hembras de lemúridos comienza en la vida adulta.
Este temprano inicio determina muy precozmente el numero
definitivo de células germinales que ira desapareciendo
hasta agotarse a medida que la hembra envejece. En el
macho esta línea celular no se agota por la
existencia de células madres (stem cells),
presentes en el tubo seminífero. Comprende
las mismas etapas que la espermatogénesis.
La mayor diferencia entre ovo y espermatogénesis
radica en que esta ultima, la meiosis se inicia en la
pubertad.
La ovogénesis comienza poco después que
las CGP colonizan la cresta genital. Estas células
denominadas ahora monocitos primordiales, pierden su actividad
migratoria y su riqueza en fosfatasa alcalina y en
lípidos. Luego empiezan a dividirse mitoticamente para dar
origen a la ovogonias.
Prenatal o período de proliferación:
empieza cuando las células germinales primordiales
colonizan el lugar donde se originaran los ovarios, allí
las células realizan divisiones mitóticas sucesivas
y forman las ovogonias, éstas se encuentra en el interior
de los folículos, al tercer mes de embarazo aumentan de
tamaño y duplican sus cromosomas y originan los ovocitos
de primer orden, se inicia la primera división
meiótica.

Postnatal o período de crecimiento: comienza
desde el nacimiento hasta el inicio de la pubertad.
Los ovocitos de primer orden continúan en el momento en
que se detuvo la meiosis.
Madurez sexual o periodo de maduración: se inicia
desde los comienzos de la pubertad hasta el momento en que la
mujer experimenta los cambios debido a la menopausia. En la
pubertad se caracteriza por la actividad del hipotálamo y
la hipófisis, este período se lleva a cabo mediante
la meiosis produciéndose las divisiones sucesivas y
continuas que caracterizan a esta división celular. El
ovocito de primer orden (2N), entra en la división
redaccional y se forman dos células hijas: ovocitos de
segundo orden, célula haploide (N) y el corpúsculo
polar de primer orden o polocitos de primer orden, célula
haploide (N).
Ambas células entran en la segunda
división dando origen a: el ovocito de segundo orden, al
óvulo (N) y un corpúsculo polar o polocito de
primer orden (N); el corpúsculo polar de primer orden a
dos polocitos de segundo orden.
El resultado final de la ovogénesis es que se
obtienen 4 células haploides (N), el óvulo y tres
polocitos de segundo orden (N), estos no son funcionalmente aptos
por carecer de los nutrientes necesarios y en consecuencia son
absorbidos por el organismo femenino y no intervienen como
células sexuales.
PERIODO OVOGENETICO
12.1.1.OVOGONIAS Y MULTIPLICACION
OVOGONIAL
En un periodo muy corto las ovogonias se dividen
activamente originando grupos de ella que en lato porcentaje
quedan conectadas por puentes citoplasmáticos. Estos
puentes son el resultado de una citocinesis incompleta. Se han
observado que aquellas que se dividen con citocinesis completa y
que, por lo tanto, están separadas de las vecinas, no
presentan alteraciones a nivel citoplasmático(vacuolas),
siendo posiblemente ellas las que entran en meiosis.
12.1.2. CELULAS SOMATICAS Y DIFERENCIACION
OVARICA
En la etapa de gónada indiferenciada, el futuro
ovario es igual al futuro testículo. En esta etapa la
ovogonias están ubicadas preferentemente en la
región blastema gonadal. La teoría clásica
considera que las células somáticas se originan a
partir del epitelio germinativo, el que invadiría el
mesenquima subyacente.
Las células del blastema gonadal y del mesenquima
ovárico originan las tecas interna y externa del
folículo ovárico.
Una vez que las células germinas quedan incluidas
en los cordones sexuales, el ovario muestra estos cordones
dispuestos como una red tridimensional formada por células
por las células germinales y por células
somáticas. ( pregranulosicas), rodeadas ambas por aun
membrana basal.
12.2. MEIOSIS Y PROFASE MEIOTICA
La meiosis es un proceso cuya mecánica es similar
en ambos sexos: sin embargo, en la ovogénesis
es el largo y discontinuo, dependiendo de variaciones
cronológicas de la especie.
La meiosis se inicia con una larga profase (PM) al final
de la cual ya no se observa ovogonias, ni ovocitos en zigoteno
paquiteno. Solamente existen células "en reposo"
denominadas ovocitos primarios, en los que la meiosis se ha
detenido, sea en estado diploteno o dictioteno.
Por el aspecto que tiene el núcleo del cito I, se
llama vesícula germinativa. Dicho ovocito esta rodeado de
un epitelio discontinuo de células somáticas
pregranulosicas, recibiendo este conjunto el nombre del
folículo primordial.
El periodo que media entre el paso de una
ovogonia a ovocito I se denomina "periodo
ovogenetico", el que se continua con el llamado periodo
folículo – genético.
Se dice que un ovario se ha diferenciado cuando las
células germinales allí presentes entran PM. Al
terminar este proceso los cordones sexuales predominantes en las
estructura ovárica en ese momento van desapareciendo. Ello
se debe a que las células del tejido conectivo
ovárico s introducen el espesor de estos cordones, los
segmentan y dejan ovocitos o grupos de los, aislados entre si.
Así se originan los llamados "nidos de ovocitos", los que
ubican se preferencia en la corteza del futuro ovario. Los
ovocitos, aislados o en grupos, están rodeados
inicialmente por un epitelio discontinuo de células
somáticas.
12.3. Función de las Hormonas
Esteroideas
La acción coordinada entre las gonadotrofinas y
los esteroides es crucial para el normal desarrollo de los
folículos ováricos, la ovulación y la
formación del cuerpo lúteo, como se
describió previamente. Ahora veamos cómo los
esteroides participan directamente en la
regulación de la función ovárica.
Andrógenos
Las células tecales producen
andrógenos, principalmente androstenediona, y testosterona
en menor proporción. Los andrógenos atraviesan la
membrana basal y ejercen efectos paracrinos sobre las
células granulosas vecinas: afectan su crecimiento,
estimulan la producción de progesterona, y además
son el substrato de la P450arom, que los convierte en
estrógenos, principalmente 17ß-estradiol. La FSH
regula la actividad de la P450arom y por ende la síntesis
de estrógenos. Tradicionalmente se ha creído que
los andrógenos inducen atresia folicular (35). Sin
embargo, ahora se conoce que ese efecto sólo se observa en
los roedores.
Estrógenos
En la mujer ingrávida los
folículos ováricos en crecimiento constituyen la
mayor fuente de estrógenos. Los estrógenos actuan
en varios tejidos donde ejercen una función endocrina
obligatoria en el normal funcionamiento del sistema reproductivo
femenino. Además, ejercen una función autocrina,
debido a que afectan a las células granulosas donde son
sintetizados, estimulando su proliferación y aumentando el
número de R- FSH y R-LH.
La acción de los estrógenos es mediada por
dos distintos subtipos de receptores nucleares, el
receptor ? (ER?) y el receptor ß(ERß). Estos
receptores son factores de transcripción que median los
efectos genómicos o transcripcionales de los
estrógenos. Los genes que expresan ambos receptores han
sido identificados en los ovarios de varios
mamíferos, incluyendo los primates. El
ERß parece ser la forma predominante en las células
granulosas, por lo que se especula que media acciones autocrinas
durante la foliculogénesis. La expresión de los
receptores de estrógenos estaría regulada por las
hormonas gonadotróficas, en particular la LH, a
través de un mecanismo dependiente del segundo mensajero
Camp.
El efecto de los estrógenos en el ovario
varía durante la foliculogénesis y es diferente
según la especie y el modelo de estudio. En los roedores
estimulan el desarrollo folicular y la proliferación de
las células granulosas .En el cerdo forman parte del
complejo de hormonas que sostienen el cuerpo
lúteo.
Corticosteroides
La función de los corticosteroides adrenales en
la respuesta fisiológica al stress está bien
establecida, aunque los efectos observados en el ovario son
aparentemente divergentes. Se ha propuesto que los
corticosteroides mediarían los efectos adversos del stress
sobre la fertilidad. Estos actuarían
indirectamente, afectando el eje
hipotálamo-hipófisis, y/o directamente sobre el
ovario inhibiendo el crecimiento folicular, la producción
de esteroides y la ovulación .Por otro lado, se han
descripto efectos beneficiosos sobre la maduración de los
oocitos y sobre la función de las células
granulosas.
La dexametasona, un análogo semisintético
del cortisol, tiene efecto potenciadores sobre la
expresión de P450scc e incrementaría la
síntesis de progesterona en las células
granulosas.
Evidencias clínicas soportan el concepto que los
corticosteroides afectan directamente la función
ovárica donde tendrían un efecto anti-inflamatorio
local. Se sabe que los niveles de cortisol en el líquido
folicular se incrementan luego de la descarga ovulatoria de LH y
que las células granulosas expresan los receptores para
los glucorticoides (GR)
12.4. FOLICULOGENESIS O PERIODO
FOLICULOGENICO.
Los folículos constituyen la estructura funcional
fundamental del ovario, siguiendo unos procesos de desarrollo
hasta alcanzar el folículo maduro, que se romperá
en la ovulación :
a) Folículos preantrales :
Folículo primordial : Formados
prenatalmente, no existen más allá de los 6 meses
de vida postnatal. Está constituido por el ovocito
primario rodeado de una única capa de células de la
granulosa aplanadas. Su evolución es independiente de la
gonadotrofinas. Folículo primario : Aumenta el volumen del
ovocito y las células epiteliales adquieren una
morfología cúbica, produciendo MPS, que originan un
halo translúcido alrededor del ovocito conocido como zona
pelúcida , atravesada por procesos citoplasmáticos
de las células de la granulosa (gap junctions) , que la
mantienen en contacto íntimo con el ovocito.
Folículo secundario (120 &µ):
Proliferan las células de la granulosa formando varia
capas y uniéndose entre ellas mediante gaps . En las
áreas en que se pierde la unión entre las
células de la granulosa se forman unas lagunas conocidas
como cuerpos de Call-Exner, que son previa a la formación
del antro por su confluencia. Se diferencian e hipertrofian las
células tecales : las internas al final del estadio
primario están separadas de la granulosa por una membrana
basal impermeable (membrana de Slavjansky , de 500 ó1000
Å) y las externas formadas por compresión del
estroma circundante ante la expansión
folicular.
La granulosa desarrolla receptores para FSH,
estrógenos y andrógenos. Con la teca el
folículo adquiere un suministro sanguíneo (1
ó 2 arteriolas que acaban en una red capilar adyacente a
la membrana basal) y las células tecales desarrollan
receptores para la LH.
Estos folículos parecen abrirse paso hacia la
superficie del ovario a través de un penacho cónico
que se forma en un polo de la teca (cono de Strassmann)
.
b) Folículo antral, terciario o madurante : La
coalescencia de los cuerpos de Call-
Exner conducen a la formación del antro folicular
, inicialmente semilunar, desplazando a las células de la
granulosa que, rodeando el ovocito, permanecen íntegras
formando el cumulus oophorus (cúmulo prolígero). El
líquido folicular está lleno de esteroides,
péptidos, proteinglicanos, etc. de los que se duda si
tienen alguna función o son mero desecho de la granulosa y
teca. Alcanza unas 1000 &µ.
c) Folículo de Graaf, ovulatorio o maduro :
Alcanza unos 15 mm, su desarrollo máximo. El cumulus
oophorus está arrinconado y constituido por la corona
radiada (células de la granulosa que envuelven al
ovocito), zona pelúcida (microvillis en el espacio
perivitelino), membrana vitelina (membrana del ovocito),
vesícula germinal (citoplasma del ovocito) y mancha
germinal (núcleo del ovocito) .
12.5. VARIACIONES CITOLOGICAS DURANTE LA EVOLUCION DE
LAS CELUALS GERMINALES.
CELULAS GERMIANLES PRIMORDIALES
La observación de las PGC al microscopio
electrónico de transmisión (MET) y microscopio
electrónico de barrido (MEB), permite apreciar su forma
redondeada, y un diámetro que oscila entre 15 y 20 mm. El
núcleo, esférico, ocupa una posición
excéntrica y contiene una cromatina granular muy fina y un
número variable de nucléolos. El retículo
endoplásmico rugoso es abundante, al igual que los
polirribosomas y los ribosomas libres. Cerca del núcleo se
observan mitocondrias esféricas con crestas lamelares,
así como un complejo de Golgi pequeño.
Asociada con el núcleo y con las mitocondrias, se
encuentra una inclusión citoplasmática muy
particular, propia de las PGC -tanto femeninas como masculinas-
llamada nuage (del francés: nube). La nuage está
constituida por masas electrodensas esféricas formadas por
material fibroso o granular, muy semejantes a los gránulos
polares ricos en ARN, característicos del plasma de las
PGC en invertebrados y vertebrados no mamíferos. El
significado funcional de esta inclusión se relaciona con
aumento en la actividad mitótica de las PGC y con la
determinación de la línea celular germinal a partir
de las células somáticas del epiblasto.
Las reservas energéticas de las PGC están
representadas por depósitos de glicógeno y
gotas de lípidos, necesarios para la
migración hacia los pliegues gonadales. También se
encuentran unos pocos microfilamentos, centríolos y
microtúbulos, así como áreas focales de
contacto estrecho con las células somáticas
vecinas.
GONOCITOS
Se diferencian de la células germinales
primordiales por la desaparición de las inclusiones
lipídicas y por la existencia de un retículo
endoplasmatico vesiculosos, poco desarrollado. El material
nuclear de estas células manifiesta un metabolismo muy
pobre.
OVOGONIAS
Su tamaño al del monocito de origen es inferior.
Son de forma esférica u oval. Generalmente están
unidas por puentes citoplasmáticos, lo que le da un
carácter sincisial. El núcleo es muy voluminoso,
siempre mas de la mitad del volumen celular. La cromatina
generalmente difusa. Los ribosomas son mas abundantes y
están distribuidos homogéneamente en el citoplasma.
Las mitocondrias muy escasas y de pequeño tamaño,
son pobres en crestas mitocondriales,
OVOCITO PRIMARIO
Luego de duplicar su ADN entran en la profase I. Estos
ovocitos I, rodeados por una capa de células planas, se
denominan folículos primordiales.
Su forma generalmente es esférica de
tamaño mayor a las ovogonias.el crecimiento ovocitario
esta acompañado de cambios muy importantes: desarrollo de
microvellosidades, producción de elementos golgianos y
aparición de gránulos corticales.
REFERENCIAS
BIBLIOGRAFICAS
– Alberts, B.; Bray, D.; Lewis, J., eds
(1996). Biología Molecular de la Célula, 3
Ed. Ediciones Omega, S.A. Barcelona.
– Arrau, E.J.,Bustos O.,Hoecker S.,Ramos
C.A.(1981).Biología de la Reproducción
Animal. Edit. Andres Bello. Chile.
– Baker.J.J.W. Garland.A. (1970).
Biología e Investigación Científica .
.Fondo Educativo
Interamericano.México
– Díaz A., Rojas N., Merzon G.,
Martínez A. (2001) Biología 2000. McGraw
Hill Interamericana de Venezuela S.A.
Caracas.
– Feliú Z. Tineo A. (2002)
Biología II año. E.M.D.P. Ediciones CO – BO.
Caracas.
– Fried. G.(1990) Biología Schaum .
MacGraw-Hill.Interamericana de México.S.A.
México.
– HALE DW (1996) Mammalian spermatogenesis.
In: VILLAR, HO (ed) Advances in Genome Biology. New York: JAI
Press Inc. pp: 249-304
– Hesham Al-Inany, M.D.
Gametogénesis y Fertilización. Gynaecology &
Obstetrics dept. Kasr El-Aini hospital, Cairo University,
Egypt.
–
http://www.kalipedia.com/ciencias-vida/tema/fases-
ovogenesis.html?x1=20070417klpcnavid_224.Kes&x=20070417klpcnavid_225
.Kes
– Mazparrote, S. (1999) Ciencias
Biológicas. 2º año de Ciencias.
Editorial Biosfera S.A. Caracas..
– Mazparrote, S. (1999 Biología
9º año: Editorial Biosfera S.A. Caracas.
– Pascual,J. (1993). Diccionario Hola
Ciencias: Susaeta Ediciones. Medellín.
Colombia.
– Rodríguez, A. y Rodríguez
C.(2000). Educación para la Salud: Editorial Romor.
Caracas.
– Ruíz. A. (1999) Biología
9º grado Editorial Triángulo. Caracas.
– Teixeira J., (2003) Ciencias
Biológicas 2. Ciclo Diversificado Librería
Editorial Salesiana S.A, Caracas.
–
http://staff.um.edu.mt/acus1/GAMETOGENESIS.htm
– Tellez G., Leal J., Bohorquez C.(1995)
Biología Aplicada. Editorial MacGraw- Hill.Latinoamericana
S.A. Bogotá.
– WASSARMAN PM, LITSCHER ES (1995)
Sperm-egg recognition mechanisms in mammals. Curr Topics Dev Biol
30: 1-19 .
–
http://www.sld.cu/galerias/pdf/sitios/histologia/matcomprepromasculino.pdf.
–
http://74.125.45.104/search?q=cache:yU1MwN1QocYJ:www.tdr.cesca.es/TDX/
TDR_UM/TESIS/AVAILABLE/TDR-1124105-133538//egr03de10.pdf+ovogonias&hl=es&ct=clnk&cd=55&gl=pe
Autor:
Paola Alvan Millones
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |