Clasificación
taxonómica.
;
Alveolata;
Apicomplexa;
Coccidia;
Eimeriida;
Eimeriidae;
Eimeria
Ciclo de
vida
Dependiendo de la especie, el ciclo de vida
es de 4 a 7 días y la diseminación se
efectúa por medio de heces, cama, polvo, escarabajos
(Alphytobius spp), moscas y otros fomites, dentro y fuera
de la granja. La coccidiosis puede dar lugar a un considerable
índice de infecciones subclínicas con diarrea y, a
veces, anemia,
trayendo como consecuencia una disminución de las tasas de
crecimiento y producción y un aumento de la
mortalidad
El número de ooquistes en la cama puede variar de
acuerdo con las condiciones climáticas, prácticas
de manejo, edad del ave y la droga
anticoccidial utilizada. Se puede constatar que las aves vivas
transportan varios estadios del parásito, permaneciendo a
veces como portadores por largos períodos
18.
En las deyecciones del huésped infectado se
eliminan los ooquistes u oocistos no esporulados, que bajo
condiciones adecuadas de humedad, sombra, presencia de oxígeno
y temperatura no
inferiores a 10°C y no superiores a 50°C sobreviven y
esporulan fácilmente dando lugar al occisto, quien
contiene 4 pequeños quistes (esporoccistos) que a su vez
contienen 2 células
infectivas o esporozoitos.
Cuando ha concluido la esporulación los occistos son
resistentes al medio y los esporozoitos son inmediatamente
infectivos para próximos huéspedes apropiados que
lo ingieran 19. Una
vez ingerido el occisto, el reconocimiento, ataque a los
componentes superficiales, fuerte unión y
penetración de la célula
hospedera se facilitan por la existencia del complejo
apical 20.
Alrededor del esporozoito se forma una vacuola donde se inician
múltiples divisiones asexuales (merogonia) durante las
cuales se forman de 2 a 100 000 merozoitos según la
especie 21.
Ya maduros los merozoitos rompen y neutralizan a las
células hospederas y migran a otras nuevas e inician una
nueva merogonia (se piensa que cada especie de Eimeria
está programada genéticamente para un número
determinado de generaciones de merozoitos de manera que el
proceso no
continúa indefinidamente, este número varía
en las pocas especies que se le conoce de 2-4)
22.
La última generación de merozoitos penetra
las células epiteliales para desarrollar la gametogonia.
La gran mayoría de los merozoitos formará
macrogametocitos, mientras que los restantes formarán
microgametocitos, ambos sufrirán múltiples
divisiones, y finalmente se obtendrán miles de
móviles y biflagelados microgametos. Cuando estos maduran
abandonan sus células hospederas y salen para penetrar
células que contengan macrogametos para producir la
fertilización.
Rápidamente tiende a formarse alrededor del
cigoto una delicada membrana y dos tipos de elementos formadores
de barrera se desarrollan dentro del citoplasma, los cuales
migran hacia la superficie de la membrana con la cual se funden
para formar una barrera resistente y cuando esta se ha formado
completamente el occisto rompe la célula
hospedera y sale en las heces. El mecanismo por el cual los
merozoitos regulan la formación de macro o
microgametocitos, así como la forma en que los
microgametocitos encuentran a las células hospederas que
contiene los macrogametocitos y los detalles del proceso de
fertilización no se han dilucidado aún
23, 24.
Esquema 1: Ciclo de las Eimerias spp
25.

Especificidad
La especificidad de hospedador es una
característica muy importante. Cada especie aviar sufre la
infección por unas determinadas especies de
Eimeria. Para vacunar contra Coccidiosis a una determinada
especie aviar se necesitan vacunas
elaboradas con un antígeno que contenga las especies de
Eimeria que con especificidad de hospedador son
patógenas para esa especie, porque hay una especificidad
inmunológica que hace que la protección sea
específica para cada especie. En la actualidad, solo
están disponibles vacunas para prevenir la Coccidiosis en
aves de la especie Gallus domesticus, pollos y gallinas,
con distinta cantidad de especies de Eimeria si la vacuna
es para reproductoras o para pollos, por su diferente ciclo de
vida. Las diferentes especies de Eimeria de las aves
también tienen una especificidad en la presentación
de las lesiones, ya que afectan a zonas determinadas del
intestino. Es la especificidad de localización, y esto
permite, junto con las características de las lesiones, la
posibilidad de un diagnóstico rápido por el
clínico en el campo.
Como se aprecia en la Figura 1, la presencia de
coccidias en el aparato digestivo
de las aves abarca la totalidad de los órganos encargados
de la absorción y procesamiento de las materias
alimenticias para el ave, tanto que se pone en riesgo permanente
el desarrollo
armónico de los animales y, en
consecuencia, se deteriora su productividad y
rendimiento.

Figura 1:
Localización de Eimerias spp en el tracto
digestivo 18
Las especies de Eimeria que afectan a pollos y
gallinas son siete: E. acervulina, E. brunetti, E. maxima, E.
necatrix, E. praecox, E. tenella, E. mitis.Luego que ha
pasado un brote con una especie de coccidia, el lote de aves
desarrolla resistencia a la
especie a que estuvo expuesto, pero siguen siendo susceptibles a
otras especies infectivas. Esto significa que el lote de aves
puede sufrir varios brotes de coccidiosis, cada uno causado por
una especie de coccidia diferente 26.
El parásito "in vivo" manifiesta una alta
especificidad lo que le ofrece facilidades para completar su
ciclo de vida y producir la infección. Las Eimeria
spp. con algunas excepciones manifiestan selectividad,
además por sistema de
órganos, órganos, tejidos y
localización dentro de la célula
27, 28.
La especificidad por el sitio a invadir es tal que se
comprobó que una administración del Eimeria a
través de una inyección endovenosa, intramuscular o
intraperitonial, producía la infección en el mismo
sitio que cuando se administra oralmente 29.
Las causas por las que cada especie de Eimeria
invade un área determinada y no otra, no han sido
fiablemente dilucidadas aunque se tienen varias hipótesis que plantean que esta
especificidad se debe a:
- El período de tiempo que
se requiera para que aparezca el esporozoito a partir del
occisto. Por ejemplo las especies de Eimeria que tienen
un gran período de esporulación tienden a invadir
la parte baja de los intestinos 30. - La presencia de determinadas moléculas en la
superficie de las células hospederas a lo largo de los
intestinos condicionará una mayor afinidad de un
área sobre otra 31.
Proceso de
invasión
Se han hecho algunos aportes con relación al
proceso de invasión de las células del hospedero
por parte de las Eimeria spp, se dijo:
- Que la introducción en la célula
hospedera ocurría a través de una fagocitosis
pasiva 32,
33. - Que los movimientos circulares y deslizantes del
esporozoito eran los que le posibilitaban reconocer, unirse y
finalmente penetrar la célula hospedera
34,
35.
Tiene su no menor importancia el hecho de que la
célula hospedera cuente con moléculas en su
superficie que podrían actuar como receptores, o productos
metabólicos que atraerían al parásito, todo
lo cual posibilitaría el reconocimiento de dicha
célula por el parásito 36.
Interacciones con
otras enfermedades.
Ha sido realizado un pormenorizado estudio sobre las
experiencias realizadas con varias enfermedades de naturaleza
bacteriana, viral y tóxica asociadas con la
coccidiosis 37. Un ejemplo de ello es la
enteritis necrótica con mortalidad elevada cuando la
presencia de clostridios precedió a la inoculación
de E. acervulina. Una significativa ocurrencia de
enteritis ulcerativa en pollos fue igualmente causada por E.
coli y E. brunetti en condiciones de campo
18.
En otros estudios, se constató el incremento de
la población de Salmonella typhimurium
en ciegos e hígado por el concurso de E. tenella,
al ser administrada 5 días consecutivos posteriors a la
inoculación de coccidias. Se observaron daños muy
serios en la mucosa del ciego cuando las inoculaciones fueron
simultáneas. Al referirse a procesos
virales, algunos autores comentan los estudios acerca de la
interferencia en el desarrollo de la inmunidad en pollos
expuestos al virus de la
Enfermedad de Marek 18,
37. En igual forma se explica el caso de la
Enfermedad de Gumboro. Cuando se expone a las aves a una
infección previa por E. tenella, se produce
hemorragia precoz con mayor mortalidad e incremento del score de
lesions. Además de estas manifestaciones, el virus reduce
el grado de protección a subsecuentes enfrentamientos a
coccidiosis cecal.
Anteriores estudios trataron el problema de la interacción entre coccidiosis y reovirus y
apreciaron una gran caída en la ganancia de peso a causa
del ataque conjunto del virus y E. acervulina, en
contraste con el efecto de cada enfermedad por separado. Dentro
de estos estudios se observó también, como
interacción, el incremento de edemas por la
asociación de los agentes etiológicos respectivos,
a comparación de lo que se observó al infectar a
las aves individualmente con cada agente 18.
Investigaciones adicionales sobre esta materia han
mostrado que los efectos adversos sobre ganancia de peso y
pigmento del plasma son mayores cuando la coccidiosis y las
micotoxicosis obran simultáneamente que cuando lo hacen en
forma no asociada . Se ha observado también que la
interacción de aflatoxina con E. tenella eleva la
mortalidad a un 60% 38.
A pesar del tiempo transcurrido, se desconoce el
mecanismo o mecanismos mediante los cuales la coccidiosis
interactúa con otras enfermedades de las aves. Se
sospecha, claro está, de una participación del
sistema inmunológico afectado o de los deterioros que
sufre el ave en el consumo,
absorción, digestión y metabolismo de
sus alimentos, lo que
deviene en una merma de sus defensas orgánicas. Se hace
necesario entonces realizar suficientes investigaciones
para encontrar la manera de reducir o eliminar los efectos
adversos de estas interacciones 18.
Control de coccidiosis por inmunidad.
El proceso de inmunidad celular se pone en marcha cuando
los ooquistes penetran en el intestino de las aves receptoras.
Los esporozoitos liberados se multiplican por división
asexual endógena de forma exponencial. Como respuesta a
esta agresión se estimulan los mecanismos de defensa del
hospedador. La inmunidad se establece después de
producirse un determinado número de lesiones y el tiempo
que tarda en alcanzarse un estado de
protección depende del número de ooquistes cantidad
de la dosis inmunizante, en el caso de las vacunas y de la edad
de los pollos.
Los antígenos que ponen en marcha el proceso
inmunitario son proteínas
de la membrana de esporozoitos y merozoitos, diferentes en cada
especie, que movilizan los mecanismos de defensa primaria. Los
esporozoitos con mayor poder
inmunógeno son los de primeras generaciones.
También tienen un pequeño papel como
antígenos en algunas especies de Eimeria, los micro
y macrogametocitos de la fase sexual del ciclo.
La respuesta inmunológica se produce en las aves
después de la primera semana de edad. Cuando se ha
establecido, actúa de la siguiente forma:
desenquistamiento de esporozoitos, freno de la penetración
de esporozoitos en las células del epitelio intestinal,
neutralización de esporozoitos en la luz intestinal y
cierta acción
del Complemento sobre los esporozoitos. Cuando el bloqueo del
parásito no es total hay una segunda barrera de
actuación en la primera generación de esquizontes.
La inmunidad humoral no juega un papel importante para luchar
contra la coccidiosis. Hay IgG, IgM y IgA en el suero de pollos
una semana después de actuar el antígeno,
alcanzando, al mes el máximo nivel. Los anticuerpos tienen
una relativa importancia, ya que los pollos sin bolsa de Fabricio
adquieren inmunidad frente a coccidiosis 39.
Grandes avances se han tenido en el
conocimiento de la respuesta inmune ante la infección
por coccidias. La infección con una especie de
Eimeria induce una inmunidad protectora exquisitamente
especifica para el parásito en particular. Un amplio de
inoculaciones de ooquistes se requiere para generar una respuesta
inmune contra Eimeria spp, aunque existen algunas
excepciones como E. máxima es altamente
inmunogenica y requiere un pequeño número de
ooquistes para generar una respuesta inmune casi completa. El
estadio endógeno precoz del ciclo de vida del
parásito es considerado más inmunogénico que
el estadio sexual tardio 40. La inmunidad no previene la
invasión de esporozoitos, pero si el desarrollo de
ellos 41.
E. tenella, provoca una respuesta
inmune que reduce los efectos de la invasión por
organismos homólogos 42,
43, la
reducción en la invasión es alrededor de un 50-65%
en comparación con las aves no inmunizadas
41, 44. Esto se cree es aparentemente un
fenómeno no específico porque la inmunidad hacia
especies de Eimeria produce una gran reducción por
parte de organismos heterológos como ocurría con
los homólogos 42,
45.
Inmunidad de mucosas
La IgG de los pollos es el anticuerpo (Ac) dominante
después de la inmunización secundaria y se
transfiere, desde la gallina ponedora al saco vitelino del huevo,
de forma similar a la transferencia placentaria de la IgG en los
mamíferos.

. Figura 2: La
IgA secretada por las células plasmáticas se une a
los receptores en la superficie interior de las células
epiteliales intestinales, después pasa a su interior y
luego dentro de las vesículas, a la superficie del
intestino y a la luz intestinal 46.
El sistema de la inmunidad de mucosas esta compuesta por
tejido linfoide asociado a mucosas (MALT) de fosas nasales,
órganos bronquiales, glándulas mamarias, tracto
genital y tejido linfoide asociado a intestino (GALT)
47.
- MALT y GALT
El papel más importante de MALT es destruir los
patógenos en su puerta de entrada para prevenir la
diseminación sistémica de la infección. Esta
función
es llevada a cabo de diferentes formas. Barreras no
específicas como las secreciones gástricas,
lisozimas, sales biliares, peristalsis y la competencia por
la flora microbiana nativa son importantes componentes de la
primera línea de defensa de MALT 48, 49.

Figura 3: Barreras
intestinales frente a la infección
50.
Las funciones
digestivas, la flora intestinal, la barrera mucosa y su
integridad y la respuesta inmune son fundamentales en la
prevención de las enfermedades entéricas.
Además de estos componentes mencionados en la figura 3,
los componentes del saco vitelino son básicos para la
defensa del pollito en el momento del nacimiento.
El mecanismo especifico para la eliminación de
patógenos nocivos involucran la interacción entre
la inmunidad humoral y la celular; aunque los Acs juegan un menor
rol en la resistencia e inmunidad a la coccidia
51, 52. Sin embargo, aves infectadas con
Eimeria producen IgA, IgM e IgY en la circulación y
en las secreciones mucosales y su habilidad para limitar la
infección es mínima. La IgY circulante e IgA biliar
especificas para coccidias son detectadas usualmente una semana
después de la inoculación de los ooquistes. Los
niveles muestran un pico que persiste por dos meses
53. Un pico de IgM
se encontró en suero a los 17 días pos
infección con Eimeria tenellae IgA biliar fue
detectada siete días pos infección con E.
tenellay E. acervulina no existiendo en la
infección secundaria. Aunque no existen evidencias
directas, IgA puede atacar la superficie de las coccidias y
prevenir la unión al epitelio, por bloqueo directo,
cambios comformacionales, impedimento estérico y/o
reducción de la motilidad. Se ha demostrado que altas
concentraciones de IgA se correspondieron con bajo número
de esporozoitos en el lumen intestinal de pollos infectados
sugiriendo un rol inhibitorio de este Ac 54. Por otra parte se ha mostrado
que IgA especifica para E .tenella puede ser detectada en
contenido cecal pero no en suero o bilis 55. También se observó
que la producción de IgA especifica fue siempre mayor en
áreas del intestino parasitadas en cualquier otra parte de
los pollos infectados con E. acervulina y E.
tenella 56.
El GALT es un tejido multiestrato continuamente expuesto
a Ag de los alimentos, de la flora microbiana normal y a
patógenos ingeridos. Más de la mitad del total de
linfocitos del MALT están contenidos dentro del GALT.
Histológicamente el estrato externo del GALT consiste en
células epiteliales y linfocitos situados encima del
basamento de la membrana, debajo de la cual se encuentra la
lámina propia (LP) que también contiene linfocitos,
además la submucosa. En pollos existe una gran variedad de
órganos linfoides especializados como las placas de payer
(PPs), tonsilas cecales y bolsa de Fabricio encargada del
establecimiento y mantenimiento
del repertorio de linfocitos B periféricos sobre todo en edades tempranas.
Tipos de células como las epiteliales, linfoides,
presentadoras de antígeno (Ag), así como las
natural killer (NK) están involucradas en el GALT para la
defensa contra patógenos 57. Otros tipos de células
incluyen macrófagos, fibroblastos, células
dendríticas entre otras. Todas ellas secretan y responden
a citoquinas siendo la
comunicación celular bidireccional.
- Células epiteliales intestinales
(IECs)
Su función primaria es absorber nutrientes
esenciales para la digestión de las comidas, además
de esto constituyen la mayor parte de la 1ra
línea de defensa contra patógenos ingeridos ya que
la mucosa de los intestinos es la más larga área de
contacto del cuerpo con el medio externo. IECs expresa el
complejo mayor de histocompatibilidad tipo II (MHC II)
58 y ha sido
observado que son malas presentadoras de antigenos (APCs) para la
activación de células T CD4 +, sin embargo pueden
procesar polipéptidos solubles y activar células T
CD8 + 47,
59. IECs produce
importantes citoquinas incluyendo IL1, IL6, IL8, factor de
necrosis tumoral (TNFµ ), inmunomoduladores, prostaglandinas
entre otros 60-63.
En la coccidiosis aviar la información es limitada acerca de la
función de IECs en la inmunidad intestinal a Eimeria
spp, aun cuando ellas están consideradas como las
1rias del hospedero utilizadas para el desarrollo
intracelular.
- Linfocitos intraepiteliales intestinales
(IELs)
Los linfocitos intestinales están presentes en
dos compartimentos anatómicos, el epitelio (IELs) y la
lamina propia (LPLs) 64. Los
leucocitos intestinales de los pollos contienen 80 % de
linfocitos, 10-15 % de monocitos, aproximadamente 5 % de
células mononucleares y menos del 1-5 de polimorfos
nucleares y células plasmáticas
64.
En pollos infectados con E. acervulina y E.
tenella se ha demostrado una diferente contribución de
células T CD4 + y CD8 + a la resistencia e inmunidad. En
pollos infectados con E. acervulina, la depleción
de células CD4 + no aumenta significativamente la
producción de ooquistes. Estos resultados sugieren que
estas células no son significativamente efectoras de la
resistencia e inmunidad para E. acervulina
65. Por otro lado
la depleción de células T CD4 + en infecciones por
E. tenella resultó en un aumento de la salida de
ooquistes en la infección primaria; pero no en la
secundaria, indicando que CD4 + son importantes efectores de la
resistencia a la infeccion primaria de E.
tenella.
La depleción de células T CD8 + disminuye
significativamente la salida de ooquistes durante la
infección primaria de ambas especies E. acervulina
y E. tenella. Esto sugiere que este efecto es debido a la
reducción del número de estas células que
sirven como transportadoras de esporozoitos. La importancia de
las células T CD8 + en la inmunidad protectora de la
infección por E. tenella es sugerida por la
observación de un aumento transitorio de
estas células en sangre
periférica durante la infección primaria
65, 66.
- Macrófagos
Los macrófagos mucosales fagocitan esporozoitos
durante infecciones con Eimeria spp, pero esta actividad
no parece aumentar la actividad de las aves
67. Los
macrófagos tienen solo la función de transportar
los esporozoitos 68. La activación de estas
células es fuente de secreción de prostaglandinas y
citoquinas que regulan la actividad de linfocitos y otros
macrófagos.
- Natural Killer (NK)
En adición los receptores de células T
(TCR+) a
b y a d
, una tercera célula mediadora de la inmunidad
intestinal son las células Killer que representan una
clase de
linfocitos citotóxicos 69-71. Las células NK carecen
de memoria
inmunológica y su actividad citotóxica no esta
restringida por moléculas MHC.
Sus efectos están mediados por varios mecanismos,
incluyendo la secreción de las citoquininas
inmunoregulatorias, lisis de células parasitadas,
interacción con células T 72.
Existen estudios que encuentran que células NK
murinas no juegan un mayor rol en la defensa contra Eimeria
sp. Cuando ratones Balb/c fueron tratados con Acs
para disminuir las células NK, no mostró cambios en
el curso de la infección por coccidias, sin embargo se
detecto un pico de actividad de células NK,
correspondiendo con una intensa respuesta inmune
73 y un
considerable desprendimiento de citoquinas
74 y IELs de pollos
contienen subpoblaciones de células NK que juegan un
determinado papel en la coccidiosis que puede ser por la
citolisis de células infectadas o como fuente
de IFN-γ 75. Otros autores reportaron que la
actividad citotóxica de NK fue drásticamente
suprimida en pollos infectados con E. tenella
76.
- Linfocitos de la lámina propia
(LPLs)
Los LPLs son predominantemente células T
activadas con menor cantidad de células B y células
plasmáticas. Investigadores han mostrado que
células CD4+ fueron encontradas exclusivamente
en la LP mientras que células CD8+ fueron
encontradas en el epitelio intestinal y en el epitelio de la
LP 77. La
mayoría de las IgA (> 80%) son originadas en sangre
periférica y migran a LP 78, 79. Otros autores reportaron que
los linfocitos IgA+ se acumulaban en la porción apical de
la LP en sitios de desarrollo de coccidias, mientras que
linfocitos IgG + fueron más numerosos en la base de la
LP 80.
De las Ig acumuladas en los sitios de desarrollo del
parásito en la LP, la mayoría fueron IgA 95 %, IgG+
(3,6 %) y IgM (1.4 %).
- Placas de Peyer
Son nódulos de tejido linfoide en la submucosa
del intestino cubiertos por un epitelio especializado el cual
provee un importante sitio para el contacto entre el Ag
foráneo y tejido linfoide. Las placas de peyer contienen
células B, células T y APCs. Estas juegan un rol
importante en la iniciación de la respuesta inmune
regulando la producción de IgA.
- Tonsilas Cecales
Es el tejido linfoide más grande del ciego y
constituye la colección mayor de GALT en los pollos. Los
linfocitos en las tonsilas consisten en 45-55 % de células
B y 35 % de células T, estando involucradas ambas en la
producción de Ac y las funciones mediadoras de la
inmunidad 81. Anatómicamente las
tonsilas están localizadas en la unión iliocecal y
no esta presente en el momento de ampollar, pero se desarrollan
poco después.
Muchos autores han sugerido que las tonsilas cecales
estan involucradas en la respuesta inmune a Eimeria sp.
Por ejemplo, durante la infección de E. tenella
aumenta el número de linfocitos, y se han encontrado
nódulos linfoides prominentes en la base de las tonsilas
aproximadamente tres centímetros de la unión
iliocecal, acumulando agregados densos de linfocitos que
contienen tejido linfoide disperso irregularmente y centros
germinales 82,
83.
Patogénesis de la coccidiosis
La mayoría de los parásitos entericos
incluyendo coccidias invaden la mucosa intestinal e inducen
daños de cierto grado en las células epiteliales e
inflamación 84. La manifestación mas
común de la coccidiosis es la diarrea
85. Los merontes
gamontos y ooquistes causan marcadas alteraciones en IELs del
hospedero en un corto periodo de tiempo, incluyendo
distorsión, ruptura, separación de células
adyacentes y formación de costra. Un extenso daño
conduce así a la diarrea, deshidratación, perdida
de peso, prolapso rectal, disentería y serios signos
clínicos al huésped pudiendo causar la
mortalidad 86. En general animales
jóvenes son más susceptibles a la coccidiosis y
manifiestan mayor cantidad de signos de enfermedad, mientras los
pollos adultos son relativamente resistentes a la
infección 56. Los animales jóvenes que
se recuperan de la coccidiosis pueden compensar en cierto modo el
crecimiento perdido pero su desarrollo potencial queda
severamente comprometido. La magnitud de los signos
clínicos resultantes de la infección por Eimeria
spp es modulada por factores genéticos del hospedero.
Por ejemplo en dos cepas diferentes de pollos de una misma raza,
SC (B2B2) y TK
(B15B21), se ha visto diferentes grados de
enfermedad con E. tenella y E. acervulina. En general
pollos SC son mas resistentes que los TK a la coccidiosis. Estas
dos cepas de pollos han sido extensamente estudiadas para el
entendimiento de la inmunidad adquirida en la coccidiosis
87.
Síntomas
Ante un brote de coccidiosis las aves se tornan
pálidas, débiles, tienden a acurrucarse, consumen
menos alimento y agua, tienen
diarrea, se pueden deshidratar, sufren pérdida en la
ganancia de peso y las ponedoras experimentan una baja en la
postura.
La coccidiosis cecal puede producir excrementos con
sangre, y anemia, que muchas veces es seguida de la muerte. La
coccidiosis intestinal no es aguda y es de naturaleza más
crónica. Produce menos mortalidad que la forma
cecal 88.

Figura 4: Apariencia
de pollos enfermos con Coccidiosis Aviar
Coccidiosis Subclínica
A raíz de la introducción de
coccidiostatos en la alimentación de las
aves, se ha ido logrando efectos cada vez menos severos de esta
enfermedad. Sin embargo, con frecuencia se habla de coccidiosis
subclínica que, en virtud de su evolución silenciosa, es fuente de
desastrosas consecuencias económicas, por su responsabilidad en el decrecimiento de la ganancia
de peso y aumento del índice de conversión
37.
En lo que respecta a entidades nosológicas,
varios autores han observado que la coccidiosis subclínica
puede asociarse con problemas de
patas en las aves o bien con deficiencias de Se o cuadros
tóxicos por Cu y Co y también puede exacerbar los
efectos de un incremento de Mg, Cu o Zn en la dieta, por la
participación de E. acervulina
18.
Diagnóstico
Como ya se ha mencionado son siete especies las que
afectan a los pollos E. acervulina, E. brunetti, E. maxima, E.
mitis, E. necatrix, E. preacox y E. tenella
1. Cada especie se
desarrola en una particular localización dentro del tracto
digestivo.
Es común encontrar al menos seis especies E.
acervulina, E. tenella, E. brunetti, E. mitis y E.
preacox en las camadas de aves de hasta las primeras seis
semanas 89 .
Cinco de las especies E. acervulina, E. brunetti, E.
máxima, E. necatrix y E. tenella son conocidas
e identificadas con relativa facilidad debido a que producen
lesiones graves con sus características propias con una
patogenicidad con un rango desde moderado hasta severo. Por otra
parte la E. preacox y E. mitisno matan los pollos o
producen lesiones patognomónicas y finalmente han sido
consideradas como benignas. Pero como quiera que sea infecciones
experimentales dan como resultados enteritis, diarreas y
reducen la eficiencia del
alimento indicando esto que estas dos especies ciertamente pueden
causar perdidas comerciales y por lo tanto necesitan ser
controladas 2.
Generalmente las cinco más patógenas
pueden ser diferenciadas en los hospederos por sus signos
clínicos, características particulares de la
lesión, sitio de infección en el intestino,
período prepatente, tamaño del ooquiste y morfología
del estado intracelular.
En los años 1990 se demostró que sondas de
RNA y DNA pueden ser usadas para identificar especies
individuales a través de características de
restricción de fragmentos patógenos
90. Técnicas
de DNA recombinante han sido usadas para discriminar diferentes
cepas de E. tenella 91 y desarrollar marcadores para
cepas resistentes a las drogas
92. El PCR ha sido
usado para diferenciar 6 especies de Eimeria además
en estudios epidemiológicos de Coccidiosis aviar
93.
Los métodos
antes mencionados son muy efectivos para la detección de
estos parásitos pero resultan muy costosos como para poder
utilizarlos en el diagnóstico diario en condiciones de
producción. Por lo que el diagnóstico de las
infecciones en el campo puede ser alcanzado por
consideración de la apariencia de síntomas
clínicos, lesiones macroscópicas y
microscópicas, detección y recuento de ooquistes,
estado de las heces, grado de las lesiones en el intestino y la
localización de las partes afectadas.
Patología de las especies de Eimeria de
las aves
Las lesiones que produce la infección dependen de
la especie de Eimeriasde su patogenicidad y la etapa en
que está la enfermedad. La cecal, puede producir
hinchazón de las bolsas cecales, que se llenan con sangre
fresca. En una etapa posterior, el ciego está lleno de un
material de aspecto caseoso, manchado con cantidades variables de
sangre. Las lesiones de la coccidiosis intestinal varían
desde una enteritis bastante suave hasta una de tipo
necrótico o hemorrágico
La severidad de la infección con cada especie de
Eimeria depende de varios factores que
incluyen:
- Edad del hospedero
- Número de oocistos ingeridos
- Edad de los oocistos ingeridos
- Susceptibilidad innata del hospedero
- Estado inmune del hospedero
- Virulencia innata de la cepa de
Eimeria

Figura 5:
Localización de las principales especies de
Eimerias en las aves 39
La distribución de lesiones dentro del
intestino es generalmente un rasgo distintivo de cada especie,
aunque un solapamiento en los sitios preferidos de desarrollo
puede ocurrir. Además, la magnitud de esparcimiento dentro
de los incrementos en el intestino con ingestión creciente
de oocistos; por ejemplo: lesiones grandes de E. necatrix
producidas durante infecciones naturales puede ser encontradas
desde el duodeno hasta la cloaca. Hay límites a
la utilidad del
diagnóstico por consideración de la apariencia de
lesiones, como las infecciones con dos especies del pollo (E.
mitis y E. praecox) que no causan cambios grandes detectables
y al menos otras dos especies (E. necatrix y E. maxima)
inducen lesiones que dependen del número de oocistos
ingeridos y la virulencia de las cepas individuales, dando una
apariencia similar 94.
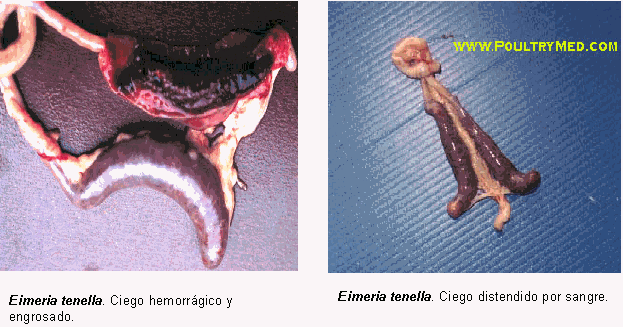
Figura 6: Lesiones causadas por
Eimeria spp 95.

Tratamiento.
Los esfuerzos de las Ciencias
Veterinarias para frenar y controlar los riesgos de la
coccidiosis han sido permanentes y de frecuente
renovación, en vista de la extraordinaria capacidad y/o
resistencia que sus agentes etiológicos han demostrado
frente a los procedimientos
que se aplicaron contra ellos 96. Una forma genérica de
apreciar este punto de vista contempla:
- Medidas de orden terapéutico, tales como el
empleo de
coccidiostatos (tratamiento preventivo) y coccidicidas
(tratamiento curativo) - Medidas de orden inmunológico, es decir,
vacunaciones (tratamiento premunitario).
La Tabla 2 presenta la diversidad de fármacos que
se viene empleando con propósitos preventivos o curativos.
Cabe destacar dentro de esta información el respectivo
mecanismo de acción ya que, en base a él, se
condiciona los diferentes progamas de rotación de
coccidiostaticos, materia de la cual se hablará más
tarde.
Es igualmente importante prestar atención al período de retiro de
estos fármacos, porque permite afirmar la ausencia total o
la presencia mínima de residuos en los tejidos,
considerando las actuales exigencias del mercado de
exportación sobre este punto y una mayor
seguridad para el
consumidor.
Tabla 2: Drogas
anticoccidiales aspectos de su farmacodinamia y
posología 18.

Clasificación
de los anticoccidianos
Los anticoccidiales atendiendo a su modo de
acción sobre Coccidias se dividen en:
coccidiostáticos, que son aquellos que inhiben
o detienen el desarrollo de los parásitos, y en
coccidicidas que son los que los destruyen
97 . Los coccidiostáticos al no
eliminar totalmente los occistos permiten el desarrollo de
inmunidad en el ave, utilizándose especialmente en los
reemplazos dedicados a la reproducción o las ponedoras,
mientras que los coccidicidas, al destruir los parásitos
inhiben el desarrollo de la inmunidad y se usan actualmente
solamente en los pollos de ceba, debido a su corto
ciclo de vida. Además se plantea que los compuestos
anticoccidiales no actúan siempre sobre todas las especies
de Coccidias y que además pueden inducir
resistencia después de un uso
continuado, por lo que se trabaja
constantemente en la búsqueda de nuevos
compuestos 98.
Los programas de
control de
coccidiosis en la producción de pollo se han enfocado
principalmente en el uso de aditivos anticoccidianos.
Los datos de investigación del mercado indican que
aproximadamente el 98% de los pollos a nivel mundial reciben un
anticoccidiano como un ingrediente estándar de su
alimento. Tres principales clases de compuestos han sido
desarrollados como anticoccidianos:
- Ionóforos poliéteres
- Químicos sintéticos
- Sapogeninas Esteroidales
Estas clases de productos tienen beneficios y
debilidades en los programas comerciales de
coccidiosis.
Ionóforos
Se han identificado más de 100 ionóforos.
Hasta ahora, solo 6 han sido desarrollados como aditivos
anticoccidianos sin vislumbrarse alguna nueva alternativa a corto
plazo. Monensina, lasolacida, salinomicina, narasina,
maduramicina y semduramicina son todos producidos por fermentación. Recientemente, alrededor del
80% de todos los alimentos medicados y que son administrados al
pollo de engorda contiene un ionóforo. Cada ionóforo tiene
fuerzas y debilidades específicas con respecto a su
espectro de acción y potencia.
La monensina, narasina y salinomicina son
intrínsecamente más eficaces en contra de E.
acervulina, pero es menos eficaz en el control de E.
tenella y E. maxima. La lasolacida y maduramicina son
más eficaces contra E. tenella, pero menos
efectivas contra E. acervulina. Los niveles
de uso de los ionóforos comercializados caen
dentro del rango de 25-100 ppm con excepción del
Maduramycin que tiene eficacia a 5
ppm 7.
El uso constante de ionóforos en las mismas
instalaciones resulta en el desarrollo de resistencia coccidiana
a esta clase de compuestos. A causa del modo de acción
similar de todos los ionóforos, la resistencia cruzada
entre ellos es muy alta. La efectividad de un producto
anticoccidiano está relacionada en gran forma a la
longitud de tiempo que un compuesto ha sido usado en la misma
instalación. A causa de la similitud del modo de
acción de todos los ionóforos y el alto nivel de
resistencia cruzada, los programas de rotación de un
ionóforo a otro tienen solo un nivel mínimo de
prevención. Los ionóforos han sido las
drogas que últimamente se han utilizado
con mejores resultados en la prevención y
terapéutica de la coccidiosis. Desde el
descubrimiento de los ionóforos, estas han sido las drogas
más utilizadas en la lucha contra la coccidiosis
en las aves
Mecanismo de acción.
- Los ionóforos tienen la propiedad de
acomplejar y transportar cationes iónicos deprotonados,
el transporte
específico de Na +, K+ y
Ca++ a través de la membrana guarda estrecha
relación con sus efectos farmacológicos. Los
ionóforos carboxílicos (Monensin, Lasalocid
y Salinomycin) intervienen en un intercambio
eléctricamente neutro de cationes por protones, sensible
al pH e
independiente del potencial de membrana, sin la
participación de los canales iónicos, o sea,
mediante una difusión dosis dependiente. Las descargas
electroquímicas del gradiente a través de la
membrana intramitocondrial causan una disminución de la
producción de ATP y un aumento de su utilización,
lo que finalmente conduce a la muerte
celular 99.
A diferencia de los compuestos químicos, los
coccidicidas ionóforos se caracterizan por:
- Afectar el equilibrio
de los iones de los parásitos, produciendo una
disminución de la energía y muerte. - No ofrecen un 100% de eficacia, sino que permiten
cierta "filtración". - Permiten que se desarrollen algunas lesiones leves
(de 1 a 2+). - Minimizan la presión
selectiva, al permitir que sobrevivan las cepas
sensibles. - Inducen el desarrollo de una respuesta inmune como
resultado de la presencia continúa de algunos
parásitos. - Reducen la proliferación de cepas
resistentes. - La eficacia de los distintos compuestos varía
de acuerdo con las especies de Eimeria
presentes. - Pueden ser tóxico o alterar la productividad
cuando se lo administra a niveles elevados o en otras
especies - No se requiere un período de retiro (aunque la
productividad puede aumentar en algunos casos)
100.
Tabla 3. Principales coccidiostáticos
ionóforos 39

Monensina y Lasolacid: Se
utilizan generalmente en pollos Broiler con una dosis de 68-113
g/ton de pienso, para la prevención de la coccidiosis
causada por E. tenella, E. brunetti, E. necatrix, E.
maxima, E. mivati, E. acervulina
101.
Salinomisina: Tiene acción
coccidiostática en la primera fase del desarrollo de los
coccidios en el intestino del pollo. Impide la penetración
de los esporozítos en el epitelio intestinal, actúa
sobre esporozítos, trofozoitos y la primera
generación de esquizontes. No debe administrarse a
gallinas ponedoras comerciales 102.
Maduramisina: Se usa profilácticamente en
pollos parrilleros, para las especies siguientes: E.
tenella, E. brunetti, E. necatrix, E. maxima, E.
mivati. Los animales tratados con Tiamulina no deben ser
alimentados con piensos que contengan MADURACYN 500 desde 7
días antes hasta 7 días después del
tratamiento, pues pueden sufrir profunda depresión
del crecimiento 102.
Nicarbazina Narasina: Para la prevención y
control de la Coccidiosis Aviar en pollos parrilleros causada por
las siguientes especies: E. tenella, E. brunetti, E.
mivati, E. necatrix, E. acervulina y E. maxima. No
utilizar en aves en postura ni en recria o mezclar con otros
coccidiostáticos 102.
Compuestos Sintéticos
Diferentes compuestos sintéticos con estructuras
químicas completamente diferentes se han desarrollado para
usarse como anticoccidianos. Cada compuesto tiene su propia
fuerza y
debilidad con respecto a la eficacia en el desarrollo de
resistencia coccidiana.
Los coccidicidas químicos presentan las
siguientes características típicas:
- Ofrecen una eficacia cercana al 100% en el control de
la coccidia al interrumpir el ciclo de vida del
parásito - Pueden actuar como coccidicida o
coccidiostático, dependiendo del químico en
particular, la especie de Eimeria predominante, y la
vía metabólica que afecta la droga. - Proporcionan un muy buen control de lesiones debido a
la interrupción efectiva del ciclo de vida del
parásito. - Su potente eficacia disminuye el nivel de
desafío coccidial. - Ejercen una fuerte presión selectiva sobre la
población coccidial, lo que genera una
proliferación de cepas resistentes. - Limitan la respuesta inmune del huésped
inducida por el parásito. - Su uso prolongado permite el desarrollo de
resistencia anticoccidial. - Pueden ser tóxico o alterar la productividad
cuando se lo administra a niveles elevados o a otras
especies. - Por lo general requieren un período de
retiro antes de la faena 100
- Por lo general requieren un período de
Tabla 4. Principales coccidostaticos
químicos 39.

Amprolium: Efectivo contra E.
tenella y E. brunetti, pero no muy efectivo contra
otras especies de Eimeria. El producto ha sido
comercializado durante 35 años y ha sido ampliamente
utilizado en Cuba, con
resultados aceptables al valorar la viabilidad y la
ganancia de peso 103. Sin embargo, Morch (1978)
en Dinamarca ha planteado que luego de un uso
prolongado, este producto demostró una actividad
pobre 104.
También en estudios realizados en Cuba este
producto resultó ser menos efectivo que la Salinomicina
cuando se midió la diferencia entre la ganancia de peso y
la conversión 105.
Toltrazuril: Es una triacinona
simétrica de la casa comercial Bayer se señala que
es un coccidicida eficaz que no afecta la inmunidad
106.
Diclazuril: Es muy potente, de amplio espectro
contra todas las especies de Eimeria. Tiene bajo potencial
para el desarrollo de resistencia para E. tenella y E.
maxima; sin embargo la inducción de la resistencia para E.
acervulina se desarrollaron más rápido que para
las otras especies 107.
Halofuginona: Este producto ha mostrado una
buena efectividad como coccidicida tanto en infecciones
experimentales frente a E. tenella, E. acervulina, E. maxima,
E. necatrix, E. mitis y E. brunetti,
como en infecciones naturales en pollos de
ceba 107,
108. Sin embargo
otros autores plantean que es altamente efectivo contra E.
acervulina, pero poca efectividad contra E. tenella.
Debe ser usado con cuidado especial debido a su potencial para un
rápido desarrollo de resistencia. De igual manera
al enfrentar este producto con diferentes especies de
Coccidias se ha encontrado que más del 90% de
las especies fueron sensibles al mismo 109.
Por otra parte, en Cuba se han realizado
experimentos
para comprobar la efectividad de la Halofuginona frente a
cepas de E. tenella y una mezcla de cepas intestinales,
habiéndose obtenido buenos resultados al evaluar la
viabilidad y peso final 110. En experimentos de campo
se comprobó que al utilizarlo solo o combinado
con el Zoalene, utilizando a este último
primero y la Halofuginona al final obtuvieron
también buenos resultados, pero al evaluar la
combinación Halofuginona-Zoalene se
observó que no se obtenían los
mejores resultados 111
.
Clopidol: Suministrando este
producto en el pienso a razón de 125 ppm, fue capaz
de proteger a los pollos contra infecciones experimentales
de E. tenella de 50 000 a 250 000 occistos
esporulados 112. Por otra parte obtuvieron
buenos resultados en la viabilidad y peso final al enfrentar
infecciones de E. tenella y una mezcla de cepas
intestinales 110.
Zoalene: Se ha encontrado una respuesta
pobre de los pollos que consumían este producto en el
pienso 104.
En experimentos realizados en Cuba en infecciones de E.
tenella, se observó una mortalidad del
100% 110.
Este medicamento no tiene igual efectividad frente a
diferentes especies de Coccidias, lo que ha sido
demostrado por otros autores al evaluarlo frente a E.
tenella, E. necatrix y E. brunetti 113.
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |