Definición anatómica de aparato y sistema:
Un aparato es un conjunto de órganos que contribuyen
a una función principal y que
necesariamente tienen una secuencia anatómica. Por ejemplo
aparato digestivo.
Un sistema es un conjunto de órganos y estructuras análogas que
cumplen funciones particulares e
interactúan con el fin de generar una función principal
general. Estos órganos no necesariamente tienen una
secuencia anatómica. Por ejemplo el sistema endocrino.
Definición histofuncional de aparato y
sistema:
Un aparato es un conjunto de órganos que contribuyen
a una función principal realizando cada uno funciones
similares con cierta independencia.
Ej.: aparato urinario. Este tiene como
componentes a los riñones, los uréteres, la vejiga y la
uretra. Los riñones se encargan de elaborar la orina y
transportarla hasta los uréteres, estos y la uretra cumplen
funciones netamente de transporte, la vejiga cumple
funciones de almacenamiento y transporte de
orina. Todos estos órganos cumplen sus funciones con cierta
independencia, y vemos q contribuyen a una única
función principal: la excreción de orina.
Un sistema conjunto de órganos y estructuras
análogas que cumplen funciones particulares las cuales
interactúan con el fin de lograr un objetivo común.
Ej.: sistema cardiovascular. Este
sistema tiene como elementos principales: el corazón, las arterias, los
capilares y las venas. La contracción del miocardio del
corazón permite el flujo de sangre a través de las
arterias, de aquí la sangre llega hasta los capilares de
todo el cuerpo, en donde la sangre arterial realiza intercambio
de sustancias con los tejidos del cuerpo, luego la
sangre poco oxigenada pasa a través de las venas y de
ahí al corazón nuevamente. Como vemos aquí hay
interacción entre las
funciones que cumplen cada uno de los elementos del sistema
cardiovascular, pues la función de uno depende del otro y de
las interrelaciones entre ellos. Vemos que el sistema
cardiovascular cumple diversas funciones como intercambio de
oxígeno y CO2, transporte
de hormonas y otras sustancias
inductoras, entre otras. Pero todas estas funciones van a
contribuir una función principal general u objetivo del
sistema cardiovascular, el cual es regular la presión arterial.
Elementos de un sistema aplicados a los elementos del
sistema cardiovascular:
· Entrada o insumo o impulso (input):
Aplicado al sistema cardiovascular, este elemento de entrada
está dado por la fibra muscular cardiaca, cuya
contracción permitirá el arranque del sistema
cardiovascular.
· Salida o producto o resultado
(output): Aplicado al sistema cardiovascular, el resultado o
producto está dado por una función general de este
sistema, en la que convergen todas las funciones específicas
de cada elemento del sistema, esta función es la
presión arterial.
· Procesamiento o procesador o transformador
(throughput): Aplicado al sistema cardiovascular, el proceso está dado por el
mecanismo de contracción de la musculatura cardiaca.
· Retroacción o retroalimentación o
retroinformación (feedback): Aplicado al sistema
cardiovascular, la retroalimentación está dada por el
flujo de retorno de la sangre a través de las venas.
· Ambiente: Aplicado al sistema
cardiovascular el ambiente estaría dado por
todos lo órganos y tejidos del cuerpo que son irrigados
mediante el sistema cardiovascular, es decir todo el cuerpo humano.
· Fronteras, restricciones o
limitaciones: Aplicado al sistema cardiovascular, las
fronteras pueden ser posibles enfermedades que imposibilitan el normal
funcionamiento del sistema cardiovascular.
BIBLIOGRAFÍA:
- BLOOM y FAWCETT. "Histología". 12va edic.
Edit. Interamericana Mc Graw Hill. Madrid, 1995 - www.es.wikipedia.org/wiki/Sistema
- www.monografias.com/trabajos39/politica-sistemica-estado/politica-sistemica-estado.shtml.
- www2.netexplora.com/gasco/ili260/textos/tgsbertalanffy.pdf
- www.elprisma.com/apuntes/administracion_de_empresas/teoriageneraldesistemas/
- www.es.wikipedia.org/wiki/Sistema_biol%C3%B3gico
2.
SISTEMA CIRCULATORIO:
IDENTIFICAR LOS SUBSISTEMAS Y SUS ELEMENTOS
ANATOMOHISTOLOGICOS.
Los sistemas principales de
transporte son el sistema circulatorio, en el que las sustancias
están disueltas o suspendidas en líquido y son
transportadas de una parte del cuerpo a otra por un sistema de
tubos (vasos). El sistema circulatorio incluye dos grandes
subsistemas:
Ø Sistema circulatorio sanguíneo:
Compuesto por una bomba muscular, el corazón y dos
sistemas de vasos sanguíneos.
Es el principal medio de transporte de oxigeno, dióxido de
carbono, nutrientes y productos de degradación
metabólicos, células del sistema
inmunitario y otros sistemas defensivos y mensajeros
químicos (hormonas), y muchas otras sustancias importantes
por ejemplo factores de coagulación.
Desde un punto de vista fisiológico (funcional) este esta
formado por tres sistemas circulatorios sanguíneos
principales:
o Circulación Pulmonar: transfiere sangre
desoxigenada con un alto contenido en dióxido de carbono
desde una bomba central a los pulmones y transporta sangre
reoxigenada desde los pulmones hacia el corazón.
o Circulación Sistémica: Transfiere
sangre oxigenada desde una bomba central a todos los tejidos
corporales y devuelve sangre desoxigenada desde los tejidos hacia
el corazón.
o Sistemas portal: son conductos vasculares
especializados que transportan sustancias de un lugar a otro,
pero no dependen de una bomba central. Ej. el sistema portal
hepático que transcurre entre el intestino y el
hígado.
Desde un punto de vista histológico este esta formado por
dos sistemas principales:
o Sistema Arterial: Que esta constituido por todos
aquellos vasos que salen del corazón y tienen paredes mas
gruesas, son de diámetro mas pequeño que las venas, y
además presentan diferentas considerables con las venas en
sus paredes.
o Sistema Venoso: Que esta constituido por todos
aquellos vasos que llegan al corazón y tienen paredes mas
delgadas, son de diámetro mayor que las arterias.
Ø El sistema Linfático: Que es el
encargado de drenar el liquido extracelular de los tejidos,
retornándolo al sistema circulatorio sanguíneo
después de pasar por los ganglios linfáticos. Este
sistema también participa en la absorción de nutrientes
desde el intestino.
ELEMENTOS ANATOMOHISTOLOGICOS:
Ø ARTERIAS
o Túnica Intima: es la capa mas
interna de las arterias y esta formada por:
§ Una única capa de células endoteliales
(epitelio escamoso simple) que secretan fibras colágenas
tipos II, IV y V, endotelina, oxido nítrico y factor de
Willebrand, además de tener enzimas unidas a su membrana como
la enzima conversora de angiotensina.
§ Tejido conectivo subendotelial: formado por tejido
conectivo laxo y unas cuantas células de músculo liso
dispuestas longitudinalmente.
o Túnica Media: es la capa mas
gruesa de la pared del vaso y esta formada por:
§ Capas de músculo liso dispuesto circularmente
§ Fibras elásticas orientadas circularmente
§ Colágena tipo III
§ Proteoglucanos
o Túnica Adventicia: Es la capa
mas externa de la pared del vaso y esta formada por:
§ Fibroblastos
§ Fibras de colágena tipo I
§ Fibras elásticas orientadas longitudinalmente
§ Vasa vasorum
§ Nervi vasorum
o Elástica interna: Banda delgada de
fibras elásticas dispuestas entre la túnica íntima
y la túnica media.
o Elástica externa: Banda delgada de
fibras elásticas dispuestas entre la túnica media y la
túnica adventicia.
Ø VENAS
o Túnica Intima: Es la capa mas
interna de las venas y esta formada por:
§ Una única capa de células endoteliales
(epitelio escamoso simple)
§ Lamina basal
§ Válvulas en algunas
venas
§ Tejido conectivo subendotelial
o Túnica media: la capa más
delgada de las venas y en algunas puede estar ausente y esta
formada por:
§ Músculo liso
§ Fibras colágenas
o Túnica Adventicia: Es la capa mas
externa de la pared de las venas y es mas gruesa que la
túnica media, esta formada por:
§ Fibras de músculo liso orientado
longitudinalmente
§ Células de músculo cardiaco cerca de su
entrada al corazón
§ Fibroblastos
§ Fibras colágenas
Ø VASOS LINFATICO
o Los vasos linfáticos presentan elementos
parecidos a las venas, con paredes mas delgadas y sin
separación nítida entre las capas intima, media y
adventicia. Sin embargo presentan mayor cantidad de válvulas
en su interior. La linfa circula por ellos gracias a la acción de fuerzas
externas (contracción de los músculos
esqueléticos) sobre sus paredes.
BIBLIOGRAFIA
- Histología Humana / Alan Stevens y James S. Lowe /
Segunda Edición / Editorial
Hacourt España S.A /
Madrid-España-1997. - Histología basica texto y atlas / Junquera y
Carneiro / Quinta Edicion / Editorial Masson / 2000 - Texto Atlas de histología / Leslie P. Gartner y James
L. Hiatt / Segunda edición / Editorial Mac Graw
HillInteramericana / Mexico-2002.
3.
DEFINIR E IDENTIFICAR LOS ELEMENTOS Y LAS FUNCIONES DEL SISTEMA
VASCULAR SANGUINEO Y DEL SISTEMA VASCULAR
LINFATICO.
SISTEMA VASCULAR SANGUINEO
FUNCIONES: Los vasos sanguíneos, los cuales son
conductos que están distribuidos por todo el organismo, se
encargan de transportar la sangre desde el corazón hasta los
tejidos del cuerpo y después la devuelven al
corazón. Forman un circuito cerrado con el corazón
ELEMENTOS:
ARTERIA: Es la encargada de distribuir la sangre
desde el corazón, hasta el lecho capilar por todo el cuerpo.
Forma el sistema arterial La acción cíclica bombeante
del corazón produce, en el sistema arterial, un flujo de
sangre pulsátil. Con cada contracción de los
ventrículos (sístole), la sangre es empujada dentro del
sistema arterial provocando la expansión de las paredes
arteriales; la subsiguiente contracción de la pared arterial
ayuda al mantenimiento de la
presión arterial entre los latidos ventriculares
(diástole). Esta expansión y contracción es una
función del tejido elástico de las paredes arteriales.
El flujo de sangre desde los distintos órganos y tejidos
está regulado por las variaciones en el diámetro de los
vasos distribuidores. Esta función se ve facilitada por la
disposición circunferencial del músculo liso de
las paredes de los vasos y está principalmente bajo el
control del sistema nervioso simpático y
las hormonas de la médula suprarrenal.
Las paredes de los vasos arteriales están formadas por la
estructura de tres capas,
general en el sistema circulatorio, aunque le caracteriza la
presencia de una importante elástica y capa de músculo
liso en la pared, gruesas en relación al diámetro de la
luz.
Arteria Elástica: Estas comprenden los grandes
vasos de conducción e incluyen las arterias aorta,
innominada, carótida común y subclavia junto a la
mayoría de los vasos arteriales pulmonares.
Tamaño estimado: El calibre de estos vasos va desde 1 cm.
o más.
La elasticidad de sus paredes
permiten hacer mas regular el flujo. La energía
cinética aplicada a los vasos durante la diástole se
convierte en energía potencial al dilatarse
elásticamente las paredes, de modo que en la diástole
dicha energía se entrega como energía cinética al
sistema (bomba subclavia del corazón).
Arteria Muscular o de Distribución: Las
arterias elásticas al ir gradualmente disminuyendo el
diámetro y el espesor de su pared, van dando lugar a ramas
laterales en las que las paredes contienen menos elástica y
más músculo liso. Entre éstas están las
principales ramas de distribución del árbol arterial
(arterias radial, femoral, coronaria y cerebral).La gran
mayoría de los vasos arteriales de la distribución
controlan flujos.
Arteria de Transición Y Arteria Especializada: en
la transición gradual de un tipo de arteria a otro, es a
veces difícil clasificar la región intermedia. Algunas
arterias de calibre de intermedio (por ejemplo, la arteria
poplítea y la arteria tibial) tienen paredes que se parecen
a las de las arterias mayores, mientras que algunas arterias
grandes(por ejemplo, la iliaca externa) tienen paredes que no
difieren de las de las arterias de tamaño medio. La
región de transición entre arterias elásticas y
musculares de llama a menudo, arterias de tipo mixto.
La arteria especializada esta referida a las
características que las diferencian de otras arterias debido
a la función que cumplen tales como: las arterias
coronarios que por estar sometidas a una gran presión,
tienen una pared que es mas gruesa que la de otras arterias
musculares de tamaño semejante. También los vasos
sanguíneos pulmonares que tienen su pared relativamente mas
fina, por ser la circulación pulmonar considerablemente mas
baja que la circulación sistémica dentro de la cavidad
craneal donde los vasos están protegidos de la presión
y estiramiento exteriores, las arterias durales y cerebrales
tienen paredes relativamente finas. Y tenemos mas haces
longitudinales de musculatura lisa en la túnica intima en
las arteria que están sometidas frecuentemente a doblarse
como las poplíteas o axilares, que en vasos comparables de
otras regiones del organismo.
ARTERIOLA: Son estructuras vasculares más
pequeñas con respecto a las arterias musculares. Vemos menos
células endoteliales, proporcionales a la superficie
endotelial (se reduce el lúmen). Los núcleos de
las células endoteliales hacen cierta prominencia hacia la
luz del pequeño vaso. Presentan una capa media gruesa,
muscular con 4 a 5 capas de músculo liso. Por fuera, hacia
la adventicia hallamos un tejido de sostén conformado por
abundantes fibras colágenas que delimitan claramente la
adventicia de la media.
La función principal de estos vasos arteriales es la de
control del flujo sanguíneo al territorio capilar (vasos de
resistencia periférica).
En las arteriolas pequeñas hallamos de 1 a 2 capas de
fibras musculares lisas, concéntricas con una lámina
elástica interna y una adventicia bastante desarrollada.
CAPILARES: Son vasos más pequeños, de paredes
muy delgadas. Su lúmen no suele ser mayor que 7 micrones.
Los glóbulos rojos pasan como tapones o émbolos por el
lúmen de estos pequeños vasos.
CAPILARES ESPECIALIZADOS
Capilares Continuos: Vasos el los cuales su
endotelio forma una delgada capa ininterrumpida alrededor de la
luz capilar. Se ubican en músculos, tejido nervioso y tejido
conjuntivo
Capilares Fenestrados: Las células endoteliales
poseen numerosos poros. En estos capilares encontramos verdaderos
"poros", aberturas o fenestraciones. Las fenestraciones se hallan
limitadas por la presencia de membrana basal. Estos "poros"
tienen un calibre determinado (90 nm. de diámetro). La
filtración se presenta principalmente a nivel de las
fenestraciones. La mayoría de los "poros" poseen diafragmas,
los diafragmas serían selectivos al regular el paso de
distintas sustancias. Cada diafragma tiene un diámetro
aproximado de 80 nm. Este tipo de capilares fenestrados los
encontramos en el estómago, intestino delgado, etc. La
presencia de poros determinaría que la cantidad de
vesículas pinocíticas generadas por la célula endotelial
sería menor con respecto a los capilares continuos.
Los encontramos en Glándulas endocrinas y sitios de
absorción de líquidos y metabolitos tales como
vesícula biliar, tubo digestivo.
Capilares Discontinuos o Sinusoidales (Sinusoides): A
diferencia de los capilares anteriores, este tipo de capilar son
mucho más anchos, con diámetros variables (30 – 70 micrones).
El trayecto de estos capilares es habitualmente "tortuoso",
siguiendo o adaptándose a la forma de los espacios que van
dejando las células con las cuales se relaciona
estrechamente. La pared de éstos capilares puede ser
continua o bien fenestrada. La lámina basal presente es
incompleta.
Encontramos capilares sinusoidales en el hígado,
médula ósea y bazo.
VENULA: Las vénulas miden entre 0,21 mm de
diámetro. Su estructura es similar a la de un capilar. A
este nivel sigue existiendo un gran intercambio de metabolitos
entre los tejidos y la sangre, junto con participar activamente
de los procesos inflamatorios, en
donde las vénulas post capilares juegan un rol relevante en
el tránsito de células inflamatorias (las vénulas
presentan el contacto endotelial menos denso de todo el Sistema
de vasos sanguíneos).
VENA: Forman el sistema venoso. Tiene una función
meramente de sistema colector de baja presión, siendo su
objetivo el retorno de la sangre desde la red capilar hasta el corazón. El flujo
sanguíneo en las venas es pasivo mediante un gradiente de
presión hasta el
corazón. La
musculatura esquelética, al contraerse permite que al
aplastarse sus paredes, la sangre "vuelva", pero este hecho no
sería suficiente sino se complementara con un sistema de
válvulas. Las válvulas permiten que se establezcan
compartimientos que junto a la presión ejercida por los
músculos esqueléticos la sangre vaya pasando de un
compartimiento al otro en su retorno.
Venas de Mediano Calibre. Histológicamente de
definen:
La íntima se presenta con una capa subendotelial poco
desarrollada que incluso, puede estar casi ausente.
La media se halla conformada principalmente por pequeños
haces de músculo liso mezclados con fibras
reticulares y fibras elásticas dispuestas en una fina red.
Su espesor no va mas allá de 3 – 4 capas de fibras
musculares lisas.
La adventicia se halla bastante desarrollada y está
constituida principalmente por colágena y fibras
elásticas. Este tipo de venas las encontramos en las
extremidades inferiores.
A medida que aumenta el espesor de la pared el vasa vasorum se
hace más manifiesto.
Venas de Gran Calibre (aquellas venas cercanas al
corazón). Histológicamente se definen:
La íntima se encuentra bien desarrollada con una media
con escasas fibras musculares lisas y tejido conectivo.
La media está escasamente desarrollada, con pocas fibras
musculares lisas.
La adventicia es la capa más evidente, con haces de
tejido muscular liso en haces dispuestos longitudinalmente.
Venas de calibre pequeño y mediano presentan
válvulas en su interior. Las válvulas son el resultado
de pliegues pares de la íntima a modo de semilunas. Es
así que finalmente las válvulas la conforma un tejido
fibroso elástico, recubiertas por endotelio.
SISTEMA VASCULAR LINFÁTICO
FUNCIONES: Sistema de drenaje encargado del
transporte de un líquido claro llamado linfa.
ELEMENTOS:
VASOS LINFÁTICOS: los capilares
linfáticos forman redes en el espacio hísticos cuyas
mallas son mayores que las de los capilares sanguíneos
vecinos, una característica importante de la pared
endotelial de los capilares linfáticos es su permeabilidad a
sustancias de tamaño molecular mucho mayor que las que
pueden atravesar la pared endotelial de los capilares
sanguíneos.
Los capilares linfáticos se unen entre sí para
formar troncos mayores que pasan por los ganglios linfáticos
vecinos o en ocasiones, a otros más lejanos. En última
instancia la casi totalidad de la linfa del organismo se acumula
en dos conductos: el conducto torácico y el conducto
linfático derecho o gran vena linfática derecha, que
vacían su linfa en los dos troncos venosos
braquiocefálicos izquierdo y derecho respectivamente.
GANGLIOS LINFÁTICOS: Son pequeños cuerpos
ovalados en forma de habichuela situados en el curso de los vasos
linfáticos, de forma que la linfa pasa a su través
antes de alcanzar el torrente sanguíneo. Suelen presentar
una pequeña depresión en una de sus
caras, denominado hilio, a través de la cual entran y salen
los vasos sanguíneos.
LINFOCITOS CIRCULANTES: Están siempre presentes en
el torrente sanguíneo, en la linfa y en otros líquidos
corporales, así como en los tejidos conjuntivos y
órganos linfoides especializados. Los linfocitos constituyen
varias poblaciones celulares que tienen en común su
capacidad de producir sustancias químicas, como los
anticuerpos, que pueden inactivar sustancias extrañas,
microbios y células neoplásicas cuando se producen en
el cuerpo o son introducidos en los tejidos.
4.
DIVISIONES DEL SISTEMA CIRCULATORIO EN HUMANOS.
Hay dos grandes sistemas circulatorios: el sistema
circulatorio sanguíneo y el sistema circulatorio
linfático.
a) Sistema circulatorio
sanguíneo: Principal medio de transporte de oxígeno,
dióxido de carbono, nutrientes y productos de
degradación metabólicos, células del sistema
inmunitario, y otras muchas sustancias importantes (por ejemplo:
factores de coagulación, mensajeros químicos,
hormonas).
b) Sistema circulatorio linfático:
Drena el líquido extracelular de los tejidos,
retornándolo al sistema circulatorio sanguíneo
después de pasar por los ganglios linfáticos; este
sistema también participa en la absorción de nutrientes
desde el intestino.
El sistema circulatorio sanguíneo se divide en:
(1) Circulación sistémica (Mayor). Es la
que transfiere sangre oxigenada desde una bomba central (el
corazón) a todos los tejidos corporales (sistema arterial
sistémico) y devuelve sangre poco oxigenada, con un alto
contenido en dióxido de carbono, desde los tejidos a la
bomba central (sistema venoso sistémico).
(2) Circulación pulmonar (Menor). Es la que
transfiere sangre poco oxigenada, con un alto contenido en
dióxido de carbono, desde una bomba central (el
corazón) a los pulmones (sistema arterial pulmonar) y
transfiere sangre reoxigenada desde los pulmones devuelta a la
bomba central (sistema venoso pulmonar).
(3) Los sistemas Portal. Son conductos vasculares
especializados que transportan sustancias de un alugar a otro,
pero no dependen de una bomba central. El sistema portal más
grande (sistema portal venoso hepático) discurre entre el
intestino y el hígado.
(4) La microcirculación. Compuesta por vasos
sanguíneos de pequeño calibre con unas paredes delgadas
parcialmente permeables que permiten el paso de algunos
componentes de la sangre hacia la sangre y viceversa. Se compone
de las metaarteriolas que vacían su contenido en los
capilares, y este es drenado luego a las vénulas.
COMPARACIÓN DEL SISTEMA CARDIOVASCULAR EN LOS
VERTEBRADOS SUPERIORES.
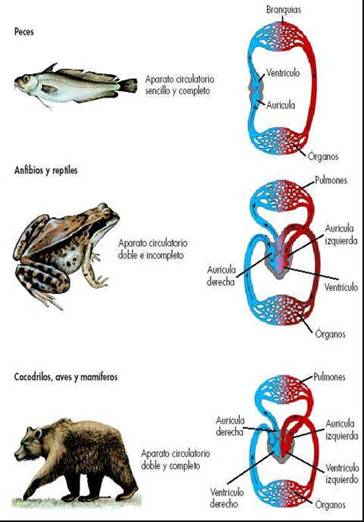
a. Peces.
§ Sistema circulatorio cerrado, simple y
completo.
§ Corazón con 4 cavidades secuenciales:
seno venoso, aurícula, ventrículo y cono. La sangre
procedente de las venas ingresa al seno venoso; al llegar al
cono, éste lo impele a la aorta ventral y a las branquias,
en estas estructuras se enriquece de oxígeno para luego
trasladarse a la aorta dorsal para su distribución en todo
el organismo; por lo tanto la sangre pasa por un solo circuito
(circulación simple), la sangre arteriosa no se mezcla con
la arterial (circulación completa).
b. Anfibios.
§ Sistema circulatorio cerrado, doble e
incompleto. Excepto en caimanes y cocodrilos.
§ Corazón de tres cámaras, con dos
receptáculos de entrada y uno de salida. En su orden: seno
venoso, aurícula derecha, ventrículo, aurícula
izquierda y cono arterioso.
§ El seno venoso se abre en la aurícula
derecha y es por donde se recibe la sangre que proviene del
cuerpo (sangre sistémica). La sangre sistémica pasa
luego a la aurícula derecha y luego al lado derecho del
ventrículo. Desde esta parte del ventrículo, la sangre
es bombeada vía arteria pulmonar hacia los pulmones. Cuando
la sangre proviene de los pulmones, esta entra a la aurícula
izquierda, vía vena pulmonar y luego al lado izquierdo del
ventrículo. Desde este punto la sangre es bombeada hacia el
cuerpo a través del cono arterioso, el cual es la parte
proximal de la aorta ventral; por lo tanto la sangre pasa por dos
circuitos (circulación
doble), la sangre venosa se mezcla con la sangre arterial a nivel
del ventrículo (circulación incompleta). Excepto en los
renacuajos: Circulación es simple y completa.
c. Reptiles.
§ Sistema circulatorio cerrado, doble e
incompleta.
§ Corazón: El tamaño, la forma,
estructura y posición de este puede variar dependiendo de la
fisiología y anatomía de cada uno de los reptiles.
Un "típico" corazón reptiliano de tortugas y escamados
tiene tres cámaras, dos aurículas (derecha e izquierda)
y un ventrículo. El ventrículo esta subdividido en tres
pequeñas cámaras o cavas (cavas arteriosa, venosa y
pulmonar, de izquierda a derecha, respectivamente). Dado que las
tres cavas no están separadas totalmente por paredes
musculares y la contracción cardiaca del ventrículo es
de una sola fase (no hay diástole ni sístole), la
sangre que esta oxigenada (sistémica) y la desoxigenada se
mezclan y salen simultáneamente a través de todos los
troncos arteriales. La sangre que se encuentra en la cava
pulmonar fluye hacia el tronco pulmonar y la sangre en el cavum
venosum fluye hacia la aorta.
§ Excepción: los Cocodrilianos, los
cuales presentan dos ventrículos bien divididos y por tanto
poseen circulación doble y completa. En cocodrilos, el
ventrículo, si esta separado completamente en dos
cámaras, izquierda y derecha. Únicamente, las dos
aortas en cocodrilos salen de cada uno de los dos
ventrículos, la aorta izquierda del ventrículo derecho
y la aorta derecha del ventrículo izquierdo. Este
patrón provee una oportunidad para que la sangre
desoxigenada se desvíe hacia los pulmones bajo
circunstancias especiales, tales como el buceo, alterando el
patrón de contracción ventricular. La sangre en
cocodrilos se puede mezclar durante un corto tiempo a través del
Agujero de Panizzae.
d. Aves y Mamíferos.
§ Circulación cerrada, doble
(circulación menor o pulmonar y circulación mayor o
sistémica) y completa.
§ Corazón con cuatro cavidades: 2
aurículas y 2 ventrículos. En vista que la
separación de aurículas y ventrículos es perfecta,
las sangre pasa dos veces por el corazón referente a cada
circuito.
Fisiología del sistema cardiovascular humano por
encima del nivel del mar.
Las características más importantes del poblador
andino son la hipertensión pulmonar y
la consiguiente hipertrofia ventricular derecha. El débito
cardiaco y la presión capilar pulmonar son normales y, por
tanto, no intervienen en el mecanismo de la hipertensión
pulmonar. El aumento de la resistencia vascular pulmonar tiene
lugar a un nivel precapilar y está relacionado con
incremento de la masa muscular de la capa media de las
pequeñas arterias pulmonares y muscularización de las
arteriolas las cuales normalmente no tienen capa muscular.
Características del sistema cardiovascular de altura:
- Arteria pulmonar es más gruesa: Su capa media contiene
gran cantidad de fibras elásticas, largas y paralelas que
le dan apariencia aórtica. - Las ramas arteriales pulmonares periféricas
(terminales) o arteriolas pulmonares a nivel del mar solo tiene
una fina capa de fibras elásticas; en las grandes alturas
mantienen su capa gruesa muscular, esta muscularización
periférica de las arteriolas aumenta su capacidad
contráctil durante el ejercicio. - Venas pulmonares con abundantes células musculares
lisas en su íntima, limitados por las capas elásticas
(interna y externa), no hay tendencia a la
arteriolización. - Las arterias terminales de la circulación pulmonar,
surgen de las arteriolas pulmonares de mediano y pequeño
calibre y se abren en el lado venoso del lecho capilar
pulmonar. Actúan como verdaderas conexiones ("bypass")
entre la circulación venosa y arterial de los pulmones. Su
rol funcional actuaría como un mecanismo compensatorio de
la hipertensión arterial. - Hay lugar a la vasocontricción pulmonar determinado
por la hipoxia y el incremento de la viscosidad sanguínea
debido a la eritrocitosis. El mecanismo intimo de la
vasoconstricción hipóxica se atribuye al efecto
perivascular de la hipoxia alveolar actuando como mediadores
locales sustancias vasoactivas (histamina, serotonina)
liberadas por los mastocitos perivasculares. - En la altura hay un incremento de la masa ventricular
derecha. El ejercicio muscular en la altura determina un
incremento de la presión pulmonar de mayor magnitud que a
nivel del mar para la misma intensidad de actividad física. Ello ocurre a pesar de que el
consumo de oxígeno y el
débito cardiaco aumentan en la misma proporción que a
nivel del mar.
BIBLIOGRAFÍA:
- Bloom. W. Fawcett. 1995. Tratado De Histología.
Doceava edición. Editorial McGraw-Hill Interamericana.
Madrid. España. - Stevens, Alan; Lowe, James. 2001. Texto Atlas De
Histología. - es.wikipedia.org/wiki/Aparato_circulatorio
- www.monografias.com/trabajos13/coraz/coraz.shtml
5. ROL DEL
CORAZÓN COMO ELEMENTO PRINCIPAL DEL SISTEMA
CARDIOVASCULAR.
El Corazón es una parte del sistema vascular muscular, de
contracción rítmica que proporciona la fuerza que impulsa la
circulación de la sangre. (Bloom Fawcett.1995.Tratado de
Histología.12ª Ed. Editorial
Interamericana)
El corazón posee un sistema propio para generar
estímulos y transmitir la
excitación producida.
(Junqueira y Carneiro.2000.Histología Básica.5ª
Ed. Editorial Masson)
El corazón como órgano de naturaleza muscular tiene el
rol de bombear la sangre hacia los
diferentes tejidos del cuerpo.
(http://preupsubiologia.googlepages.com/cardio-sangre)
El Rol del corazón es bombear la sangre a través del
sistema arterial con una presión
considerable.
(Ross, Pawlina.2005.Histología.5ª Ed.Editorial
Médica Panamericana)
El rol del corazón es actuar
prácticamente una doble bomba que suministra
la
fuerza necesaria para que la sangre circule a través de los
dos sistemas circulatorios más importantes: la
circulación pulmonar en los pulmones y la circulación
sistemática en el resto del cuerpo. La sangre primero
circula por los pulmones y posteriormente por el resto del
cuerpo.
(http://omega.ilce.edu.mx:3000/sites/ciencia/volumen1/ciencia2/37/htm/sec_8.htm)
BREVE DESCRIPCIÓN
ANATÓMICA DE LOS ÓRGANOS QUE FORMAN EL SISTEMA
CARDIOVASCULAR
El sistema cardiovascular es el conjunto de conductos
por los que circula la sangre y esta formado por:

- El corazón.
- Los vasos sanguíneos.
- Corazón:
En anatomía, el corazón es el órgano
principal del aparato circulatorio. Es un músculo estriado
hueco que actúa como una bomba aspirante e impelente, que
aspira hacia las aurículas la sangre que circula por las
venas, e la impulsa desde los ventrículos a las
arterias.
- Situación: El corazón está situado
prácticamente en medio del tórax (mediastino), entre
los dos pulmones, encima del diafragma, delante del raquis
torácico separado de las vértebras por el
esófago y la aorta, y detrás del esternón y de
los cartílagos costales. El corazón se fija en esta
situación por medio de los grandes vasos que salen y
llegan a él, y por el pericardio. - Forma y orientación: El corazón tiene
forma de pirámide triangular o cono, cuyo vértice se
dirige hacia abajo, hacia la izquierda y hacia delante, y la
base se dirige hacia la derecha, hacia arriba y un poco hacia
atrás. - Volumen y peso: El volumen del corazón
varía según el sexo y la edad.
Tradicionalmente se ha comparado el volumen del corazón
con el de un puño, pero cambia considerablemente
dependiendo de si el corazón está en sístole o
en diástole. El volumen total varía entre 500 a 800
mililitros, siendo más importante el volumen de
eyección del ventrículo izquierdo. Su peso ronda los
275 gramos en el hombre y 250 gramos en
la mujer. - Partes del corazón: El corazón se divide
en dos mitades laterales, que son el corazón derecho, en
la que circula la sangre venosa y el corazón izquierdo, en
la que circula la sangre arterial. Cada una de estas dos
mitades se subdivide en otras dos, situadas una encima de la
otra que son: la cavidad superior llamada aurícula o
atrio, y la cavidad inferior llamada ventrículo. Cada
aurícula comunica con el ventrículo por medio de un
orificio llamado orificio auriculoventricular, que
contiene una válvula derecha llamada tricúspide y una
válvula izquierda llamada mitral. Los dos corazones
están separados en toda su altura, por medio de un tabique
vertical que se llama tabique interauricular entre las
dos aurículas y tabique interventricular entre los
dos ventrículos. Por lo tanto:
- Corazón derecho: Está formado por la
aurícula derecha y el ventrículo derecho, separados
por la válvula tricüspide. - Corazón izquierdo: Está formado por la
aurícula izquierda y el ventrículo izquierdo,
separados por la válvula mitral.
- Estructura del corazón: Las capas del
corazón son de dentro afuera: el endocardio, el miocardio
y el pericardio. Entre las capas del corazón se encuentran
fibras nerviosas constituyendo el plexo cardiaco.

- Vasos Sanguíneos:
Un vaso sanguíneo es conducto hueco ramificado por
el que fluye la sangre. El conjunto de vasos sanguíneos del
cuerpo junto con el corazón forman el aparato circulatorio.
Los vasos sanguíneos se clasifican en tres
grupos:
- Venas.
- Arterias.
- Capilares.
Solo pasare a describir los vasos que se relacionan
íntimamente con el corazón para evitar la gran cantidad
de ellos por ello trataré venas y arterias y no
capilares.
a) Venas:
En anatomía una vena es un vaso sanguíneo que
conduce la sangre desde los capilares al corazón y lleva
dióxido de carbono y desechos de los organismos. El cuerpo
humano tiene más venas que arterias y su localización
exacta es mucho más variable de persona a persona que el de las
arterias.
Las venas se localizan más superficialmente que las
arterias, prácticamente por debajo de la piel, en las venas
superficiales.
Las venas de mayor calibre y que se relacionan
íntimamente con el corazón son:
Venas Cavas:
Ø Cava Superior:
" Origen, Trayecto Y
Terminación: Desde su origen que esta formado por la
unión de las venas braquiocefalicas, posteriormente al
primer cartílago costal derecho, la vena cava superior
desciende un poco oblicuamente en sentido posterior y describe
una curva cuya concavidad se adapta ala convexidad de la
porción ascendente de la aorta. Desemboca en la pared
superior del atrio derecho, ala altura de la extremidad
anterior o esternal del segundo espacio intercostal derecho.
" Dimensiones: Mide por
término medio 7 cm. de longitud y 2 cm. de
diámetro.
Ø Cava Inferior
" Origen Y
Terminación: Procede de la unión de las dos venas
iliacas comunes, anteriormente y a la derecha de la columna
lumbar, un poco inferiormente a la bifurcación de la aorta y
la altura de la porción superior de la 5ta vértebra
lumbar. La vena cava inferior atraviesa el diafragma y desemboca
en la pared inferior del atrio.
" Dimensiones: Su longitud
es por termino medio 22 cm, y su calibre es irregular y variable.
En su desembocadura la vena mida aproximadamente 3cm de
diámetro.
Ø Vena Pulmonar:
Las venas pulmonares presenta las características
morfológicas de las venas, pero llevan sangre arterial al
corazón.
Son generalmente dos para cada pulmón, una superior y
otra inferior.
" Origen: Las venas
pulmonares nacen de vénulas procedentes de la red capilar de
los alvéolos pulmonares, reciben también las venas
procedentes de las últimas ramificaciones bronquiales y de
la pleura visceral.
" Terminación: Las
venas pulmonares se dirigen medialmente y terminan en el atrio
izquierdo.
b) Arteria:
En anatomía una arteria es cada uno de los vasos
que llevan la sangre desde el corazón a las demás
partes del cuerpo.
Las arterias llevan al organismo la sangre que ha pasado a
través de la circulación pulmonar y está
enriquecida o saturada con oxígeno.
Las arterias de mayor calibre y que se ralacionan
íntimante con el corazón son:
Ø Arteria Pulmonar:
" Origen: Tiene
características externas de las arterias pero conduce a los
pulmones sangre venosa. Su origen se sitúa en el orificio
del tronco pulmonar del ventrículo derecho.
" Ramas Terminales: Las
ramas derechas e izquierda del tronco pulmonar se dirigen hacia
sus lados respectivos y alcanzan el hilio del pulmón
correspondiente.
Ø Arteria Aorta
" Origen Y
Terminación: La aorta es el tronco de origen de todas las
arterias del cuerpo. A partir del orificio aórtico del
ventrículo izquierdo. Termina a la altura de la 4ta
vértebra lumbar dividiéndose en tres ramas la arteria
sacra media y dos iliacas comunes.
" Porciones De La
Aorta:
- Porción ascendente
de la aorta
- El arco de la aorta
- Porción descendente
de la aorta
BIBLIOGRAFÍA:
- Anatomía. Keith L. Moore. 4ta edición. Editorial
Médica Panamericana. Buenos Aires – Argentina.
2002. - Anatomía Humana. H. Rouviére. Tomo I. 10ma
Edición. Masson S.A. Barcelona. 1999. - http://www.ciencia.net/VerArticulo/?idTitulo=Sistema%20cardiovascular
- http://www.texasheartinstitute.org/HIC/Anatomy_Esp/anat1_sp.cfm
7. TEJIDO
MUSCULAR ESTRIADO CARDIACO.-POBLACION CELULAR.
El tejido muscular cardiaco se forma en las paredes del
corazón y también se encuentra en las paredes de los
vasos sanguíneos principales adyacentes. Deriva de una masa
estrictamente definida del mesénquima esplácnico, el
manto mioepicárdico, cuyas células surgen del epicardio
y del miocardio.

Tejido muscular estriado cardiaco, corte longitudinal,
corazón (H-E), 1000X

Tejido muscular estriado cardiaco, corte transversal,
corazón (H-E), 1000X
MIOCITOS
La población celular del
músculo cardiaco esta formada por los Miocitos,
Que son unidades celulares separadas 80 um de longitud y
15 um de diámetro.
Los miocitos individuales se ramifican y forman conexiones
oblicuas con las bandas vecinas, formandose una compleja organización tridimensional
muy diferente del orden paralelo de las fibras cilindricas del
músculo esqueletico.
El sarcoplasma es mas abundante y tambien es mas aparente la
estriacion transversal debido a la separacion de los haces de
miofibrillas por filas de mitocondrias. Las miofibrillas
presentan una direccion divergente alrededor del
núcleo situado centralmente, rodeando una region axial
fusiforme del sarcoplasma en la que existen abundantes organelas
e inclusiones.
Las mitocondrias del músculo cardiaco presentan numerosas
crestas que muestran con frecuencia una angulacion periodica que
las hace presentar un patron en zig-zag.

El miocardio está conformado por diferentes tipos de
células y una estructura intersticial como se puede ver en
esta representación. Los miocitos son las únicas
células que responden a las diferentes señales aumentando su
tamaño (hipertrofia), las otras tienen la capacidad de
dividirse (hiperplasia).
Los miocitos presentan abundante glucogeno y aparece en
forma de particulas densas de 30 a 40 nm localizadas en areas de
sarcoplasma que quedan entre los miofilamentos y que tambien
contienen mitocondrias aunque estas particulas tambien se pueden
alinear en filas entre los propios miofilamentos.
El reticulo sarcoplasmico longitudinal es menos complejo
que el del músculo esqueletico, esta constituido por una
trama subsarcolemal de tubulos de 20 a 35 nm de
diámetro.
Antes se pensaba que las funciones de las celulas miocardicas
se limitaban a la contracción y a la exitacion o
conducción, pero en los dos ultimos decenios se ha
identificado una serie de miocitos auriculares que sintetizan y
secretan hormonas peptidicas implicadas en la regulación de
volumen de la sangre y en la composición electrolitica del
liquido extracelular . Las celulas mioendocrinas, que son celulas
especializadas que se localizan principalmente en las orejuelas
auriculares derecha e izquierda, aunque tambien se puede
encontrar de forma dispersa en otras zonas de las aurículas
y a lo largo del sistema de conducción en el tabique
ventricular. S u caracteristica que la diferencia es que presenta
granulos secretorios de elevada densidad rodeados por membrana y
de 0.3 a 0.4 um de diámetro. Estos granulos se concentran en
la parte central dl sarcoplasma que se extiende em ambas
direcciones desde los polos del núcleo aunque tambien se
pueden observar entre los miofilamentos y ocasionalmente en la
vecindad del sarcolema.
Una caracteristica exclusiva del músculo cardiaco es la
presencia de discos intercalares transversales a intervalos
regulares de la longitud de las fibras.
Estos discos intercalares presentan complejos de unión
que se encuentran en la interfase de células musculares
adyacentes. Son uniones que aparecen como líneas rectas o
muestran un aspecto en escalera. En la parte en escalera se
distinguen dos regiones. La parte transversal, que cruza la fibra
en línea recta y la parte lateral que va en paralelo a los
miofilamentos.
En los discos intercalares se encuentran tres tipos de
contactos:
![]() La fascia adherens o
La fascia adherens o
zona de adhesión
![]() mácula adherens o
mácula adherens o
desmosome
![]() uniones tipos gap (gap
uniones tipos gap (gap
juntion).
Desde el punto de vista funcional, el paso de iones permite
que las cadenas de células musculares se comportan como si
fueran un sincito (célula simple con muchos
núcleos), pues el estímulo de la contratación pasa
como si fuera una onda de una célula a otra.

Necrosis de reperfusión de fibras miocárdicas.
Partes de dos fibras separadas por un disco intercalar; a la
derecha, fibra normal; a la izquierda, fibra con bandas de
contracción, entre éstas rarefacción de
filamentos, mitocondrias mineralizadas y desplazadas en
acúmulos.
BIBLIOGRAFÍA:
- /trabajos11/teosis/teosis.shtml
- http://med.unne.edu.ar/revista/revista106/miocardiopatia_hipertrofica_revision.html
- www.fac.org.ar/ccvc/llave/c001/escudero.ph
- www.portalesmedicos.com/publicaciones/article
- www.portal.reduaz.mx/…/Carlson/Cap08/Cap08.htm
8.
TEJIDO MUSCULAR ESTRIADO.- COMUNICACIÓN
CELULAR.
Una característica del musculo cardiaco es la presencia
de líneas transversales intensamente coloreables, que
aparecen a intervalos irregulares. Estos son los discos
intercalares representan complejos de unión que se
encuentran en la interfase de las células musculares
adyacentes. Son uniones que aparecen como líneas rectas o
muestran un aspecto de forma de escalera. En las partes
escalonadas se distinguen dos regiones: la parte transversal, que
cruza la fibra en ángulo recto, y la parte lateral, paralela
a los miofilamentos. En los discos intercalares se encuentran
tres tipos de uniones principales: la zónula adherens o
fascia adherens, los desmosomas o maculae adherentes (las cuales
son uniones de anclaje) y las uniones comunicantes o
uniones de hendidura o uniones tipo gap o uniones de fisura
(uniones de comunicación).
De las cuales vamos a tratar de las uniones de
comunicación para el caso de comunicación celular.
Uniones de hendidura o uniones comunicantes:
Constituyen el elemento estructural principal del componente
lateral del disco intercalar. Las uniones de hendidura, que
tienen una gran importancia fisiológica, proveen continuidad
iónica entre las células musculares
cardíacas contiguas y de eso modo permiten que
moléculas de información pasen de una
célula a otra. Este intercambio posibilita que las fibras
musculares cardíacas o miocitos coordinen sus actividades y
se comporten como un sincitio y al mismo tiempo retengan su
integridad y su individualidad. La posición de las uniones
de hendidura en las superficies laterales del disco intercalar
las protege de las fuerzas generadas durante la
contracción.
Las uniones comunicantes son las únicas estructuras
celulares conocidas que permiten el paso directo de
moléculas de señal de una célula a otra. Una
unión de hendidura consiste en una acumulación de
poros o canales transmenbrana dispuestos muy juntos. Estas
hendiduras permiten que las células intercambien iones,
moléculas reguladoras y metabolitos pequeños a
través de los poros. La cantidad de poros en una unión
comunicante puede variar mucho, al igual que la cantidad de
uniones comunicantes entre las células contiguas.
Las uniones de hendidura están formadas por 12
subunidades de proteínas pertenecientes a
la familia de las conexinas
(proteínas que contienen cuatro hélices trasnmenbrana).
Estudios realizados sobre este tipo de uniones han permito ver
grupos de canales muy juntos, cada uno formado por dos
hemicanales llamados conexones, que están incluidos en las
membranas enfrentadas. Estos canales están formados por
pares de conexones que cruzan el espacio extracelular entre las
células contiguas. El conexón de una membrana celular
está alineado con precisión para acoplarse con un
conexón coincidente en la membrana de la celular
contigua y así, como su nombre lo indica, permite la comunicación entre las
células.
Cada conexón tiene seis subunidades simétricas de
una proteína integral de la membrana llamada conexina (Cx),
que se aparea con una proteína similar proveniente de la
otra membrana contigua. En consecuencia el canal completo
está compuesto de 12 subunidades que adoptan una
distribución circular para formar un canal cilíndrico
de 10nm de longitud y 2.8 nm de diámetro a través de la
membrana. El canal tiene un calibre de 1.5 nm.
En tejidos con células excitables eléctricamente,
las uniones de tipo gap cumplen una función importante, como
por ejemplo, el acoplamiento eléctrico entre células
nerviosas permite el transporte rápido de los potenciales de
acción de una célula a otra. También el
acoplamiento eléctrico, que se realiza en este tipo de
uniones, sincroniza las contracciones de las fibras musculares
cardíacas y fibras musculares lisas responsables del
movimiento peristáltico
del intestino.
Las uniones tipo gap no siempre están abierto sino que
alternan entre estados abiertos y cerrados. La permeabilidad de
estas uniones puede disminuir rápidamente y de forma
reversible cuando hay un descenso del pH celular o un incremento de
la concentración del Ca2+ libre
citoplasmático. Por eso a estas estructuras se las considera
como dinámicas las cuales dependen del
pH y la de regulación de la concentración de
Ca2+ en el citoplasma, lo que lleva a un cierre
temporal de los canales de las uniones tipo gap.

Canales iónicos:
Las uniones comunicantes forman poros relativamente grandes y
permisivos que resultarían desastrosos se conectan
directamente el interior de una célula con el espacio
extracelular. Por lo cual existen otras uniones más
estrechas y selectivas que se pueden abrir y cerrar. Estas
proteínas están relacionadas específicamente con
el transporte de iones inorgánicos, por lo que se denominan
canales iónicos. Estos canales tienen una eficiencia mayor que los
transportadores ya que a través de cada canal abierto pueden
pasar hasta 100 millones de iones por segundo, lo cual indica que
tiene una velocidad de 105 veces
superior a la del transporte mediado por cualquier transportador.
Los canales iónicos no pueden acoplarse a una fuente de
energía para poder realizar un transporte
activo. Por lo que el transporte que realizan es siempre pasivo.
La función de los canales iónicos es la de permitir que
algunos iones inorgánicos determinados (fundamentalmente
Na+, K+, Ca2+ o
C-) puedan difundir rápidamente a favor de su
gradiente electroquímicos a través de la bicapa
lipídica

Los canales iónicos son selectivos para el ion
transportado y fluctúan entre estados abiertos y
cerrados:
Los canales iónicos tienen dos propiedades importantes,
primero es que presentan selectividad para el ion transportado,
es decir solo transportan el ion que les corresponde no dejando
pasar a otros; segundo es que fluctúan entre estados
abiertos y cerrados, lo cual es influenciado por estímulos
que causan la abertura de los canales iónicos como por
ejemplo cambios en el voltaje (canales regulados por voltaje), un
estrés mecánico
(canales regulados mecánicamente), o la unión a un
ligando (canales regulados por ligando). El ligando puede ser
tanto un mediador extracelular, específicamente un
neurotransmisor (canales regulados por transmisor) o un mediador
intracelular, como un ion (canales regulados por iones) o un
nucleótido (canales regulados por nucleótidos).
El potencial de membrana en células animales depende principalmente
de los canales de fuga de K+ y del gradiente de
K+ a través de la membrana
plasmática:
Cuando existe una diferencia de cargas eléctricas a
ambos lados de una membrana, se establece un potencial de
membrana, debido a un ligero exceso de iones positivos sobre los
negativos en un lado y un ligero déficit del otro. Estas
diferencias de carga pueden deberse tanto a un bombeo
electrogénico activo, como a una difusión pasiva de
iones. En el caso de los animales lo que genera el potencial
eléctrico de la membrana plasmática son los movimientos
pasivos de iones. La bomba de
Na+/K+ ayuda a mantener el equilibrio osmótico a
ambos lados de la membrana de una célula animal al mantener
muy baja la concentración intracelular de
Na+. Como hay muy poco Na+ en el
interior de la célula, se acumulan otros cationes para
contrarrestar las cargas negativas de aniones fijos de la
célula como son las moléculas orgánicas cargadas
negativamente que están confinadas en el interior de la
célula. El papel equilibrador los realiza mayoritariamente
el K+, que es bombardeado activamente al interior
de la célula por la bomba de
Na+/K+ y que también puede
desplazarse hacia el interior o el exterior por canales de fuga
de K+ de la membrana plasmática.
Los canales catiónicos regulados por voltaje generen
potenciales de acción en las células excitables
eléctricamente:
La membrana plasmática de todas las células
excitables eléctricamente, como las musculares, contienen
canales catiónicos regulados por voltaje, que son los
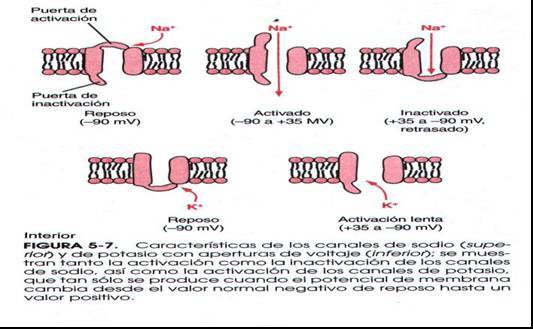
responsables de generar los potenciales de acción. Un
potencial de acción se dispara por la despolarización
de la membrana plasmática, es decir, por una variación
del potencial de membrana a un valor menos negativo. En las
células musculares un estimulo capaz de provocar la
suficiente despolarización hace que se abran unos canales de
Na+ regulados por voltaje, permitiendo que entre
en la célula una pequeña cantidad de Na+
a favor de su gradiente electroquímica. La entrada
de cargas positivas aumenta la despolarización de la
membrana, lo que provoca la abertura de mas canales de Na+ y la
entrada de mas Na+ y esto a su vez lleva a una
mayor despolarización. Este proceso continúa
autoamplificándose hasta que, en una fracción de
milisegundos, el potencial eléctrico de la membrana que ha
sido afectada pasa de su valor de – 70mV en reposo a un valor de
+50 mV, próximo al del potencial de equilibrio del
Na+. En este punto, en el que la fuerza
electroquímica que impulsa el flujo de Na+ es
casi cero, la célula llegaría a un nuevo estado de
reposo, con todos sus canales de Na+
permanentemente abiertos, si la conformación abierta del
canal fuera estable. La célula evita este espasmo
eléctrico permanente gracias a dos mecanismos que
actúan de forma concertada: la inactivación de los
canales de Na+ y la abertura de los canales de
K+ regulados por voltaje. Los canales de
Na+ tienen un mecanismo de inactivación
automático, que hace que se vuelvan a cerrar
rápidamente incluso aunque la membrana todavía
esté despolarizada. Los canales de Na+
permanecen en este estado inactivado, incapaces de volverse a
abrir, hasta unos cuantos milisegundos después de que el
potencial de membrana haya vuelto a su valor negativo inicial.
Esto nos lleva a decir que los canales de Na+
pueden encontrarse en tres estados diferentes cerrado, abierto e
inactivo.
Los canales de K+ regulados por voltaje suponen
un segundo mecanismo que, contribuye a que la membrana
plasmática activada recupere más rápidamente su
potencial negativo original y quede preparada para transmitir un
segundo impulso. Estos canales se abren, de manera que la entrada
transitoria de Na+ se ve compensada por una salida
de K+; la consecuencia de esta abertura es que la
membrana vuelve rápidamente hacia el potencial de equilibrio
del K+, incluso antes que se complete la
inactivación de los canales de Na+ .
Estos canales de K+ responden
a cambios en el potencial de membrana de la misma manera que lo
hacen los canales de Na+, pero con una
cinética ligeramente más lenta, por lo que se les
denomina canales de K+ retardados.

El registro de zona indica que cada
uno de los canales de Na+ se abre siguiendo la
ley de todo nada:
Las fibras musculares contienen muchos miles de canales de
Na+ regulados por voltaje y la corriente total que
atraviesa la membrana es la suma de las corrientes que fluyen a
través de todos ellos. Esta corriente es posible detectarla
mediante la técnica del registro de zona, un método que revolucionó
el estudio de canales iónicos. El registro de zona
indica que cada uno de los canales de Na+
regulados por voltaje se abre siguiendo la ley de todo o nada.
Los tiempos de abertura y cierre de un canal son aleatorios pero,
cuando se abre, el canal siempre tiene la misma conductancia,
permitiendo el paso de más de 1000 iones por milisegundo.
Así pues, la corriente agregada que cruza la membrana de una
célula no indica el grado de abertura de los canales sino el
número total de canales de la membrana que están
abiertos en un momento dado.
Los canales catiónicos regulados por voltaje
están relacionados evolutivamente y
estructuralmente:
Los canales de Na+, K+,
Ca2+, son regulados por voltaje y no solo
están en las células excitables eléctricamente,
sino que también en células que no son activas
eléctricamente.
En cada una de estas tres clases de canales catiónicos
existe una sorprendente diversidad estructural y funcional,
generada tanto por múltiples genes como por la
maduración alternativa de los transcritos de RNA producidos
a partir de un mismo gen. Sin embargo, las secuencias de todos
los canales conocidos de Na+,
K+, Ca2+ regulados por voltaje
muestran elevados grados de similitud, lo que sugiere que todos
ellos pertenecen a una gran superfamilia de proteínas
evolutiva y estructuralmente relacionados y que comparten muchos
de los principales motivos de diseño.
Los receptores de acetilcolina de las uniones
neuromusculares son canales catiónicos regulados por
transmisores:
Un ejemplo de un canal iónico regulado por transmisores
es el receptor de acetilcolina de las fibras musculares
esqueléticas. Este canal se abre transitoriamente por la
acetilcolina liberada por la terminal nerviosa de la unión
neuromuscular (la sinapsis química especializada entre una
neurona motora y una fibra
muscular esquelética).
El receptor de acetilcolina del musculo esquelético
está formado por cinco polipéptidos transmembrana, dos
de un tipo y tres más diferentes entre sí, codificados
por cuatro genes independientes. Cada uno de los dos
polipéptidos idénticos del pentámero contiene
centros de unión para la acetilcolina. Cuando se une dos
moléculas de acetilcolina al complejo pentamérico,
inducen un cambio conformacional que abre
el canal. Con el ligando todavía unido, el canal alterna los
estados abierto y cerrado, pero ahora tiene una probabilidad del 90% de estar
en el estado abierto. Este estado
continúa así hasta que la concentración de
acetilcolina haya disminuido lo suficiente debido a su
hidrólisis por una enzima especifica (la acetilcolina
esterasa) localizada en le unión neuromuscular. Una vez
liberado del neurotransmisor unido, el receptor de acetilcolina
vuelve a su estado de reposo inicial. Si la presencia de
acetilcolina se prolonga mucho tiempo como resultado de una
excesiva estimulación nerviosa, el canal se inactiva.

La transmisión neuromuscular implica la
activación secuencial de cinco grupos diferentes de canales
iónicos:
La importancia de los canales iónicos en las células
excitables eléctricamente se puede ilustrar mediante el
seguimiento del proceso por el cual un impulso nervioso estimula
la contracción de una fibra muscular. Esta respuesta,
aparentemente sencilla, requiere la activación secuencial de
cómo mínimo cinco grupos diferentes de canales
iónicos, en el intervalo de unos cuantos milisegundos.
1) Este proceso se inicia cuando un impulso
nervioso alcanza la terminal nerviosa y despolariza la membrana
plasmática de la terminal. La despolarización abre
transitoriamente los canales de Ca2+ regulados por
voltaje de esta membrana. Dado que la concentración de
Ca2+ fuera de la célula es 1000 veces
superior a la concentración de Ca2+ libre en
el interior, el Ca2+ entra en la terminal
nerviosa. El aumento en la concentración de
Ca2+ del citosol de la terminal nerviosa
dispara la liberación localizada de acetilcolina a la
hendidura sináptica.
2) La acetilcolina liberada se une a los
receptores de acetilcolina de la membrana plasmática de la
fibra muscular, abriendo transitoriamente los canales
iónicos asociados a ellos. Como resultado de ello, la
entrada de Na+ causa una despolarización local de la
membrana.
3) La despolarización local de la
membrana plasmática de la fibra muscular abre los canales de
Na+ regulados por voltaje de la membrana,
permitiendo la entrada de más de Na+ y
aumentando la despolarización de la membrana. Esto provoca
la abertura de los canales de Na+ regulados por
voltaje vecinos y genera una despolarización autopropagadora
(un potencial de acción) que se extiende hasta afectar a
toda la membrana plasmática.
4) la despolarización
generalizada de la membrana plasmática de la fibra muscular
activa los canales de Ca2+ regulados por voltaje
de algunas regiones especializadas de la membrana (los
túbulos transversos T).
5) Esto, a su vez, induce la abertura
transitoria de los canales de liberación de
Ca2+ presentes en una región adyacente de la
membrana del retículo sarcoplasmático y la
liberación del Ca2+ almacenado en el
retículo hacia el citosol. Este incremento repentino en la
concentración citosólica de Ca2+ es el
responsable de la contracción de las miofibrillas de la
célula muscular. No se sabe aun como es el mecanismo por el
que la activación de los canales de Ca2+
regulados por voltaje del túbulo T provoca la abertura de
los canales de liberación de Ca2+ de la
membrana del retículo sarcoplasmático, aunque es
posible que un cambio en la conformación del canal de
Ca2+ de la membrana plasmática inducido por
el voltaje pueda abrir directamente el canal de liberación
de Ca2+ del retículo sarcoplasmático
mediante un acoplamiento mecánico.

Flujo de los iones de calcio en el ciclo de
contracción – relajación cardiaca:
Patrones de desplazamiento del calcio:
No se conocen todos los detalles acera del flujo de los iones
del calcio que relacionan la contracción con la onda de
excitación, si bien es posible proponer un modelo de trabajo. Una de las hipótesis que más se
aproxima es la referente a la función elemental que tiene la
liberación de calcio a partir del SR. La mayor parte de los
datos existentes se basa en la
teoría de la
liberación de calcio inducida por calcio, que consiste en
que el SR libera iones de calcio abundantes hacia el citosol en
respuesta a la cantidad inferior que penetra en el miocito
cardiaco con cada onda de despolarización. Esta
hipótesis es también llamada
teoría de la sinapsis química.
El aumento en la concentración citosólica de calcio
concluye cuando pasa la onda de excitación, puesto que no
penetra más iones y el SR interrumpe la
liberación de calcio. Este último fenómeno se
explica a través de alguno de los mecanismos siguientes:
1) La concentración
citosólica de iones de calcio se ha elevado lo suficiente
como para inhibir el proceso de liberación del ion inducida
por calcio.
2) La liberación de este ion a
partir del SR está ligada a la apertura de los canales de
calcio, así que cuando estos últimos se cierran, el SR
suspende la liberación del ion.
3) La concentración
citosólica alta de iones de calcio activa a la bomba de
captación de calcio del SR.
4) El SR libera calcio solo durante el
potencial de acción.
Para equilibrar la pequeña cantidad de iones de calcio
que penetra en el célula cardiaca durante cada
despolarización, una cantidad similar debe abandonar
célula a través de alguno de los procesos siguientes.
En primer lugar, es posible intercambiar iones de calcio por
iones de sodio a través del intercambio Na+/Ca2+ y, en
segundo, la bomba sarcolémica de calcio que consume
ATP transfiere calcio hacia el espacio extracelular, en
contra un gradiente de concentración.
Control sarcolémico del calcio y otros iones:
Los modelos actuales de
excitación y contracción adjudican una función
fundamental a la apertura inducida por el voltaje de los canales
sarcolémicos de calcio tipo L al comienzo del proceso de
contracción. Estos canales son proteínas
macromoleculares formadores de poros que atraviesan la capa
sarcolémica doble de lípidos, proporcionando una
vía altamente selectiva para transferir iones hacia la
célula cardiaca cuando el canal se abre. Los canales
iónicos tienen dos propiedades principales: la
formación de compuertas y la penetración. Cada canal
está protegido por dos o más compuertas
hipotéticas que controlan su apertura. Los iones solo
penetran a través del canal cuando ambas compuertas se
encuentran abiertas. En el caso de los canales de sodio y calcio,
que son los mejor conocidos, la compuerta de activación se
cierra durante el potencial de reposo normal de la membrana
y se abre la compuerta de inactivación, de manera que las
compuertas de los canales son controlados por el voltaje. La
despolarización abre la compuerta de activación.
Estructura molecular de los canales de calcio tipo
L:
Existe una similitud molecular sorprendente entre los canales
de sodio y de calcio. Este hallazgo muestra una conservación de
la estructura quizá de todos los canales iónicos
controlados por voltaje, y sugiere una familia genética común. Ambos
canales contienen una subunidad alfa mayor, con cuatro
subunidades o dominios transmembrana de estructura similar.
Además, tanto los canales de sodio como los canales de
calcio contienen en su estructura global otras subunidades que
funcionan de manera menos conocida, como la subunidad beta. Cada
uno de los cuatro dominios transmembrana de la subunidad alfa
está formado por seis hélices. En cada dominio existe un segmento
helicoidal específico, llamado S4, que es rico en
aminoácidos, con carga altamente positiva y que
supuestamente constituye la localización del sensor de
voltaje.
La subunidad beta tiene el efecto de acentuar el flujo de
calcio a través de los poros de la subunidad alfa. La
estructura de aminoácidos de los poros tiene propiedades
fundamentales. Por ejemplo, los residuos de glutamato ayudan a
establecer la presencia de enlaces de calcio de afinidad
acentuada y, por tanto, la especificad del poro por el ion de
calcio.
Fosforilación de los canales de calcio:
La subunidad alfa, (que es la subunidad específica de
cada órgano) del canal sarcolémico de calcio se
fosforila en distintos sitios, ante todo en la región
terminal C. durante el estimulo adrenérgico beta, el AMP
cíclico aumenta dentro de la célula y se transfieren
grupos fosfato del ATP a la subunidad alfa. De esta manera,
las cargas eléctricas cercanas a la boca interna de los
poros vecinos se alteran para inducir cambios en la
conformación molecular de los poros, de tal suerte que
aumenta la probabilidad de apertura del canal de calcio.
Canales de calcio tipos T y L:
Los canales T (transitorios), que se abren cuando el
voltaje es más negativo, lo hacen durante periodos
cortos y no interactúan con los antagonistas
convencionales del calcio. Estos canales son supuestamente
los que producen la primera fase de la apertura del canal
de calcio.
Los canales sarcolémicos L (largos) son regulares, se
localizan en el miocardio y están implicados en la
liberación de calcio inducida por calcio. Las compuertas de
estos canales funcionan de dos maneras (modos de compuertas). El
modo uno se caracteriza por episodios cortos de apertura y el
modo dos por episodios mayores.
Sustitutos iónicos:
Durante la relajación, la bomba sarcoplásmica para
captar calcio y el sustitutivo de
Na+/Ca++ compiten por la
eliminación del calcio citosólico, y normalmente domina
la bomba de SR. Para restituir el equilibrio del calcio se activa
una serie de sustitutivos transarcolémicos, de los cuales el
principal es el sustitutivo de
Na+/Ca++. Este sustitutivo (con
peso molecular de 108 KDA) consta de 970 aminoácidos y
carece de homología con cualquier otro tipo de proteína
conocida. Ya se logró identificar un péptido inhibidor
específico ([Exchange inhibitor peptide, XIP] péptido
inhibidor del intercambio). La dirección del intercambio
iónico responde al potencial de membrana, y a la
concentración de sodio y calcio en ambos lados del
sarcolema. Puesto que los iones de sodio y calcio se intercambian
hacia adentro o afuera en respuesta al potencial de membrana,
debe existir un potencial de membrana específico, llamado
potencial de inversión o de
equilibrio, con el que los iones se distribuyen de manera tal que
puedan desplazarse con la misma facilidad en cualquier
dirección.
Importancia fisiológica del sustitutivo de
sodio-calcio:
1) Es posible que la penetración
transarcolémica de calcio durante el intercambio de modo
inverso participe en la liberación de calcio inducida por
este mismo elemento.
2) El intercambiador participa en la
restitución de los equilibrios iónicos.
3) Es probable que este sustitutivo
participe en la relación entre fuerza y frecuencia
(fenómeno de Treppe o de Bowditch). De acuerdo con la
hipótesis del retraso en la bomba de sodio, la rápida
acumulación de iones de calcio durante el estimulo
rápido del miocardio sobrepasa a la capacidad del
intercambiador de Na+/Ca++ y de la bomba de sodio para volver a
la normalidad iónica. El resultado es la acumulación de
iones de calcio dentro del SR, con incremento en la fuerza de la
contracción.
Bomba de sodio:
El sarcolema es altamente permeable al Na+ solo
cuando su canal se abre al principio de la despolarización y
también penetra durante la salida de Ca2+ por
el intercambiador de Na+/Ca2+. La mayor parte de
la entrada de Na+ a través del sarcolema debe corregirse por
la actividad de la bomba de Na+/K+, llamada también ATPasa
de Na+/K+ o simplemente bomba de Na+.
La bomba es actividad por el sodio interno o por el potasio
externo. Por cada ciclo de transporta se utiliza una
molécula de ATP. Al principio los iones se encierran dentro
de la bomba proteínica y luego salen hacia cualquiera de los
lados. Si bien existe cierta controversia respecto de la
proporción exacta de Na+ y K+ bombeada, el modelo
más aceptado es el que afirma que por cada tres Na+ que
salen entran dos K+. Durante este proceso, una carga positiva
debe de abandonar la célula. Por tanto, la bomba es
electrógena, así que también se le llama bomba
electrógena de Na+. La corriente inducida por la actividad
sostenida sostenida de la bomba aporta alrededor de -10Mv al
potencial de reposo de la membrana. Puesto que la bomba debe
extraer iones de Na+ a través del intercambio de Na+/Ca++ o
por el canal de Na+, su actividad sostenida es fundamental
para mantener el equilibrio iónico normal.


BIBLIOGRAFIA:
· Alberts
B, A. Johnson, J. Lewis, M. Raff, K. Roberts y P. Walter. 2006.
"Biología Molecular de la
Célula". 4a Edición. Editorial Omega.
Barcelona- España.
·
Braunwald. Tratado de cardiología (volumen I y II).
5a edición. Editorial MCGRAW-HILL
INTERAMERICANA. México- 2000
9.
SARCOMERA CARDIACA-
PROTEINAS CARDIACAS.
La maquinaria contráctil del músculo cardiaco esta
representada por las miofibrillas. Las mismas están
compuestas por unidades contráctiles denominadas
sarcómeras de 2,2 um de longitud y un ancho equivalente a la
miofibrilla. Con microscopio electrónico se
puede ver una estructura electro densa denominada disco Z que
separa una sarcómera de otra. Este disco se encuentra
ubicado en una región poco densa llamada la
banda I (por isotrópica) en donde solamente hay filamentos
finos. Estas bandas alternan con otras denominadas bandas A
(por anisotrópica) donde se hallan filamentos gruesos y
finos. En la parte media de las bandas A se encuentra la
banda H de menor densidad aun (donde solo hay filamentos
gruesos). Estas distintas bandas sufren variaciones
periódicas que se deben a la superposición de las
proteínas cito esqueléticas.
En la sarcómera pueden distinguirse los filamentos de
actina (filamento fino) que nacen de los discos Z, donde existe
la alfa-actinina que es la proteína que une la actina y la
titina, esta ultima es una proteína elástica (la
más grande del organismo). La titina posee dos
funciones:
ü mantiene a la miosina en su posición
y, debido a que tiene una parte elástica,
ü actúa como resorte recuperando la longitud
de la miofibrilla después de la contracción
muscular.
La miosina, proteína que forma el
filamento grueso, esta formada por dos cadenas ligeras (muy
parecidas a la calmodulina y troponina c pero que han perdido la
afinidad por el calcio), que forman la cola y dos cadenas pesadas
que formas la cabeza. Cada cadena pesada esta compuesta por
tres dominios: uno el terminal NH2, el segmento central y el
extremo terminal COOH, estos dos últimos son los que
interaccionan con la actina.
El filamento fino esta formado por actina G proteína
globular la cual se va polimerizando para formar un filamento de
actina F que luego se combinara con otro para formar el
filamento de actina.
Cada siete pares de actina G encontramos un complejo de
proteínas reguladoras formado por la troponina T que
se une a la tropomiosina, la troponina C que
es la que tiene afinidad por el calcio y la Troponina I
que tiene función inhibidora. Todas estas proteínas
tienen forma globular.
La calsecuestrina (CASQ) es la proteína mas abundante en
el interior del retículo sarcoplásmico, donde
actúa como amortiguador evitando la precipitación del
Ca2+ que se encuentra en altas concentraciones en el RS (2-10
mM), y también regulando la capacidad de liberación de
Ca2+ por el receptor de rianodina (RyR).
10. COMPARACIÓN
HISTOBIOQUIMICA DE LA SARCOMORA CARDIACA CON LA
SARCÓMERA ESQUELéTICA.
La sarcómera se define como la unidad Anatómica y
Funcional del músculo, por ser la mínima parte del
músculo capaz de realizar las funciones contracción y
relajación.
La observación de la a
través de microscopio revela que la sarcómera
tiene fibras presenta una serie de estriaciones alternantes
claras y oscuras. Las bandas claras son las bandas I
(isotrópicas), y las bandas oscuras, con su elevado
índice de refracción, son las bandas A. En la parte
central de la banda A se localiza la banda H, una región de
baja densidad óptica causada por la
ausencia de filamentos de actina. En la mitad de la zona H,
existe una región oscura, la "línea M", la cual
marca el centro del
sarcómero. La región M está formada por
estructuras proteicas filamentosas que conectan de forma cruzada
a los filamentos de miosina, manteniendo su arreglo y dando un
espaciamiento regular entre ellos. También en la línea
M se anclan los filamentos conectores. El segmento entre dos
líneas Z adyacentes se denomina sarcómera, de unas 2 a
3um de longitud, de las cuales 1.5 m corresponden a la banda A y
0.8 a la banda I. La sarcómera no sólo es una unidad
estructural, sino también la unidad contráctil
básica. En el músculo relajado en corte longitudinal
las bandas están bien marcadas. Durante la contracción,
las miofibrillas son más gruesas y las sarcómeras
más cortas, ya que la distancia entre las líneas Z se
acorta en forma progresiva. A medida que las bandas I se hacen
más cortas, los extremos de las bandas A se aproximan a las
líneas Z, hasta que al final, en contracción total, las
bandas A e I son indistinguibles; pero la longitud de la batida A
en la contracción permanece constante.
Proteínas presentes en la sarcómera
muscular:
- Actinina: proteína fijadora de actina, organiza
los filamentos finos en forma paralela y los ancla en la
línea Z. - Miomesina: proteína fijadora de miosina,
mantiene los filamentos gruesos alineados en la línea
M. - Titina: proteína larga extremadamente elastica,
que discurre paralela ala serie de filamentos y une las
terminaciones de los filamentos gruesos al disco Z. - Desmina: una proteínas de filamentos
intermedios, que forma una malla alrededor del sarcomera a la
altura de la línea Z, con lo que une estos discos entre si
y a la membrana plasmática y forma enlaces cruzados
estabilizadores entre miofibrillas vecinas. - Proteína C: proteína fijadora de miosina
que tiene la misma función que la miomesina y forma varias
franjas transversales bien definidas a cada lado de la
línea M. - Nebulina: una proteína alargada,
inelástica de 600 KDa que esta adherida a la línea Z
y transcurre paralela a los filamentos finos.

Diferencia entre una sarcomera cardiaca y una sarcomera
esquelética:
característica | T.M.esquelético | T.M.cardiaco |
forma | Cilíndrico extremos ramos. | Cilíndricos extremos ramificados |
núcleo | Multinucleado periférico | 1 a 2 núcleos disposición |
Uniones intercelulares | No | Si; células unidas por discos |
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |