Contracción | voluntaria | Involuntaria, ante estímulos nerviosos. |
velocidad | rápida | intermedia |
regeneración | Puede regenerarse (tiene células | No puede regenerarse (no tiene cél. |
La principal diferencia entre la sarcomera
esquelética y la sarcomera cardiaca es que en la sarcomera
esquelética vamos a encontrar un complejo llamado triada
compuesto por 2 cisternas terminales y un tubulo T entre las
bandas A e I; mientras que en la sarcomera cardiaca vamos a
encontrar un complejo llamado díada, compuesto por una
cisterna termina y un tubulo T a nivel de la línea Z
Diferencias entre músculo esquelético y el
músculo cardiaco:
BIBLIOGRAFÍA:
- Michael H. Ross, PhD. Texto/Atlas de Histología. 4ta
Edición. Ed.
Médica Panamericana. Argentina. 2004
- Gartner,L., Hiatt, J. Texto Atlas de Histología. 2da
edición. Mc. Graw – Hill Interamericana Editores, S.A. de
C.V. México. 2002.
- Bloom – Fawcett. Tratado de Histología. duodécima
edición. Ed. Interamericana. España. 2002.
11. CONTRACCIÓN CARDIACA, IDENTIFICAR LOS ELEMENTOS
HISTOLÓGICOS QUE LA SUSTENTAN.
ELEMENTOS HISTOLÓGICOS QUE SUSTENTAN LA
CONTRACCIÓN CARDIACA:
- El
conjunto de miofibrillas que contienen fibras de actina y
miosina, además de otras, que forman la sarcómera
cardiaca y permiten la contracción.
- Junto
a cada miofibrilla (estructuras que usan
energía) hay abundantes mitocondrias grandes (estructuras
que liberan y recapturan energía) y depósitos de
glucógeno (almacén de energía). Lo
cual asegura una contracción continua.
- Los
discos intercalares, poseen uniones celulares en sus dos
componentes ( lateral y transversal) que van a permitir:
o Formar fibras musculares cardiacas
funcionales, que los filamentos finos del sarcómero terminal
se fijen a la membrana plasmática. (Fascia adherens)
o Impedir que las células cardiacas se
separen ante la tensión de las contracciones regulares
repetidas. (Desmosomas)
o Proveer de continuidad iónica entre
las células musculares cardiacas contiguas y de ese modo
permitir que moléculas de información pasen de una
célula a otra. Además
que las fibras se comporten como un sincitio, contrayéndose
coordinadamente. (Nexos)
-
Canales de Ca2+ sensibles al voltaje en la pared del túbulo
T, son la causa de un retraso, característico y necesario
para la contracción rítmica, de unos 200 milisegundos
desde el comienzo de la despolarización en la
contracción muscular cardiaca.
- El
latido cardiaco es iniciado, regulado localmente y coordinado por
células musculares cardiacas modificadas que están
especializadas y reciben el nombre de células de
conducción cardiaca. Estas células se organizan en
nódulos (células cardionectoras nodales) y
fibras de conducción ( células cardionectoras
fasciculares) muy especializadas (Fibras de Purkinje) que
generan y transmiten con rapidez el impulso contráctil a las
diversas partes del miocardio en una secuencia precisa.

BIBLIOGRAFÍA:
- ROSS, Michel; PAWLINA, Wojciech. Histología, Texto y
Atlas color con Biología Celular y Molecular, 5ª
edición, Buenos Aires: Médica
Panamericana, 2007. - D. W. FAWCETT. Tratado de
histologia 12° edicion,
1995 editorial McGraw hill Interamericana.
12. TEJIDO MUSCULAR ESTRIADO
CARDIACO RENOVACIÓN Y/O REPARACIÓN Y/O
REGENERACIÓN. ADELANTOS CIENTÍFICOS: NUEVAS
TEORÍAS.
El músculo cardíaco no tiene, prácticamente,
capacidad de regenerar. Los daños del músculo
cardíaco se reparan por proliferación del tejido
conjuntivo, produciéndose una cicatriz.
Las estimaciones de la tasa de proliferación de los
miocardiocitos eran diversas, pero los estudios más
cuidadosos la situaban en el 0,0005%. No puede sorprender, por
tanto, que en los casos de una muerte masiva de
miocardiocitos a causa de un infarto no fatal se produzca
un tejido cicatricial rico en tejido conectivo, pero sin
capacidad contráctil. El rendimiento del corazón queda pues
inevitablemente disminuido tanto en este caso como en otros que
implican un deterioro de la pared cardiaca.
En el corazón postnatal normal, los cardiomiocitos
en mitosis son aproximadamente
![]() y se estima que
y se estima que
la perdida progresiva de cardiomiocitos es de ![]() cada año,
cada año,
lo que demuestra que a lo largo de la vida se produce un recambio
lento de los mismos a partir de células madres cardiacas y
otros tejidos.

ADELANTOS CIENTÍFICOS: NUEVAS
TEORÍAS:
Células madre cardiacas y
cardioblastos:
Ramón Muñoz-Chápuli
Catedrático del Departamento de Biología Animal,
Universidad de
Málaga
¿Cómo sería posible recuperar funcionalmente
esa parte dañada del corazón? En principio podemos
distinguir dos grandes alternativas de terapia celular que
implican, respectivamente, el implante en el corazón de
distintos tipos de células extracardiacas o el recurso a las
propias células del corazón. La primera alternativa, la
que hasta ahora se ha ensayado en humanos, es la que
tratábamos en su momento en nuestro anterior artículo
sobre la cardiomioplastia, por lo que no nos extenderemos ahora
sobre ella, salvo para señalar las novedades que desde
entonces se han producido. Esta posibilidad trata de explotar la
capacidad que tienen determinados tipos celulares de
diferenciarse en miocardiocitos para repoblar con ellos la zona
infartada. La posibilidad de obtener algunos de estos tipos
celulares del propio paciente y expandirlos en cultivo evita los
problemas de rechazo
inmunitario. Los tipos celulares que pueden utilizarse con este
fin son:
– CéLULAS MADRE
EMBRIONARIAS: son fácilmente expandibles en cultivo y se
diferencian en miocardiocitos con facilidad. Sin embargo, aparte
de las cuestiones éticas, su utilización se ve
dificultada por el hecho de ser alogénicas (provocan
rechazo) e incluso oncógenas si no se controla
cuidadosamente su estado de diferenciación
antes de la implantación en el paciente.
– CéLULAS SATéLITE DEL MÚSCULO
ESQUELéTICO (MIOBLASTOS): constituyen una población minoritaria del
músculo estriado (alrededor del 3%) pero tienen la propiedad de regenerar dicho
músculo. Fueron los primeros tipos celulares implantados en
el corazón, donde forman miotubos contráctiles. Estos
miotubos ocasionalmente se fusionan con los miocardiocitos, pero
en general permanecen eléctricamente aislados del miocardio.
Los ensayos clínicos
realizados muestran que los mioblastos implantados forman
grupos de miotubos alineados
con los miocardiocitos incluso 18 meses después de su
implantación [Menasche et a., J Am Coll Cardiol 41:1078-83
(2003)]. En general, el procedimiento implica una
mejoría en la función cardiaca, sea por la
propia capacidad contráctil de los implantes o por efectos
indirectos de los mismos. No obstante, habrá que esperar al
final de los ensayos clínicos para elaborar conclusiones
definitivas.
– CéLULAS DE LA MéDULA ÓSEA: en esta
población podemos distinguir las células madre
hematopoyéticas (HSC), las mesenquimáticas (MSC) y los
progenitores endoteliales (EPC). Estas células se han
utilizado frecuentemente en la terapia celular de la isquemia
miocárdica, y han dado lugar a una importante polémica
científica, en particular sobre su capacidad de
diferenciarse o no en miocardiocitos funcionales. En efecto, han
sido varios los estudios que ponen en tela de juicio dicha
capacidad de diferenciación, aunque casi todos coinciden en
que esta terapia es beneficiosa para el estado del paciente y
mejora el funcionamiento cardiaco. Algunos estudios señalan
que los presuntos casos de diferenciación observados
serían una consecuencia de la fusión de las
células de la médula ósea con miocardiocitos
[Nygren et al., Nat Med 10(5):494-501 (2004)], y que la mayor
parte de las células implantadas en el corazón dan
lugar a células hematopoyéticas (CD45+) pero no a
células cardiacas [Balsam et al., Nature 428:668-73 (2004)].
Otros indican que la permanencia de las células de la
médula ósea en el corazón es muy breve (alrededor
de 4 semanas), aunque insisten en la mejoría del
funcionamiento cardiaco, probablemente por una mejor
vascularización [Limbourg et al., Eur J Heart Fail 7:722-9
(2005)] o por efectos paracrinos [Kinnard et al., Circulation
109:1543-49 (2004); Gnecchi et al., Nat Med 11:367-68 (2005)]. En
sorprendente contraste con estos estudios, otros muestran
indicios de lo contrario, es decir, que las células de la
médula ósea sí se diferencian en miocardiocitos
funcionales cuando son implantados en el corazón, que
permanecen a largo plazo y que su diferenciación no se debe
a la fusión [Kajstura et al., Circ Res 96:127-37 (2005);
Eisenberg et al., Stem Cells en prensa, (2006)]. En ciertos casos
se ha señalado que no son las HSC sino las MSC las que
tienen la capacidad de diferenciación miocardiocítica
[Kawada et al., Blood, 104:3581-7 (2004)].

¿A qué se debe esta disparidad de resultados?
Probablemente a los modelos experimentales usados
(humanos o no humanos), a los criterios de selección, purificación
y expansión de las células de la médula ósea
y a la técnica utilizada para implantar las células en
el corazón (infusión vascular, inyección directa).
No obstante, lo que parece quedar claro después de numerosos
ensayos clínicos es que la función cardiaca mejora
después del implante de células de la médula
ósea, bien sea por diferenciación hacia miocardiocitos,
por diferenciación hacia células vasculares (con el
consecuente mejor riego de la región afectada) o incluso por
un efecto paracrino que mejora la supervivencia del tejido
cardiaco.
Células madre cardiacas y
cardioblastos
La gran novedad que se ha producido en estos últimos
años ha sido el descubrimiento de células residentes en
el propio corazón con capacidad para generar miocardio. En
concreto, se han descubierto
dos poblaciones celulares diferentes en el corazón que, a
pesar de su baja frecuencia, podrían desempeñar un
papel importante en la homeostasis cardiaca, es
decir, en la reposición de los miocardiocitos perdidos por
apoptosis o necrosis. Este era un tema realmente misterioso, ya
que no se conseguía «cuadrar el balance» de las
pérdidas de miocardiocitos por causas naturales. Como hemos
señalado antes, todo apunta a que sólo uno de cada
200.000 miocardiocitos humanos se encuentra en el ciclo celular
[Rubart y Field, Annu Rev Physiol 68:29-49 (2006)]. Esta cantidad
se estima como insuficiente para mantener la masa del
corazón más allá de unos pocos años, por lo
que se sospechaba que debía existir algún tipo de
mecanismo celular de reposición. Las dos poblaciones
recientemente descubiertas son:
– CéLULAS MADRE CARDIACAS. Estas células
expresan marcadores típicos de células madre (son
c-Kit+ Sca-1+) y poseen las tres características
básicas de dichas células, esto es,
auto-renovación, clonogenicidad y multipotencialidad.
Inyectadas en el corazón en modelos animales son capaces de
diferenciarse en miocardiocitos y en vasos sanguíneos
[Beltrami et al., Cell 114:763-76 (2003); Linke et al., Proc Natl
Acad Sci U S A 102:8966-71(2005)]. Estas células ya se han
aislado de corazones humanos y se han cultivado in vitro,
condiciones en las que forman «cardiosferas»
pulsátiles [Messina et al., Circ Res 95:911-21 (2004)].
– CARDIOBLASTOS O PROGENITORES CARDIACOS. Son
células muy poco abundantes en el corazón (500-600 en
el corazón de la rata recién nacida). No expresan
marcadores de células madre y sí el producto del gen homeobox
Islet-1, que se expresa durante el desarrollo del sistema nervioso y que marca los progenitores cardiacos
tempranos durante el desarrollo embrionario. Podemos
considerar a las células Isl1+ como auténticos
progenitores cardiacos que permanecen en el corazón. Pueden
ser aislados, cultivados sobre células nutricias, expandidos
y reinyectados en el corazón, donde se diferencian en
miocardiocitos [Laugwitz et al., Nature 433:647-653 (2005)].
Como ya hemos dicho, es probable que una de estas poblaciones, o
quizá ambas, contribuyan a la homeostasis celular del
corazón humano. El problema es que estos mecanismos de
regeneración son demasiado lentos para responder en un caso
catastrófico de infarto en el que mueren millones de
miocardiocitos en pocas horas. No obstante, el conocimiento de la
existencia de células madre cardiacas y cardioblastos
sí permite concebir la posibilidad de obtenerlos mediante
biopsia, expandirlos in vitro y reimplantarlos al paciente.
La terapia con células madre cardiacas o
cardioblastos plantea una posibilidad complementaria o incluso
alternativa en un futuro a la terapia con células de la
médula ósea, y no es difícil prever que si
volvemos a tratar este tema en Encuentros en la
Biología dentro de tres o cuatro años es
probable que hablemos ya de los primeros ensayos clínicos.
Pero, cuidado, no es esta la única posibilidad que existe.
Se pueden concebir al menos dos estrategias más para la
regeneración cardiaca desde el punto de vista experimental,
al tomar en consideración las propias células del
corazón:
– El estímulo de la proliferación
miocárdica. Ya hemos dicho antes que la tasa de
proliferación natural de los miocardiocitos es muy reducida,
pero también es cierto que en procesos patológicos esta
proporción se incrementa, al menos en modelos
experimentales. Las preguntas que surgen son ¿cuál es
el mecanismo molecular que promueve la proliferación de los
miocardiocitos? Y, sobre todo, ¿podemos utilizar ese
mecanismo para promover y prolongar en el tiempo esta
proliferación? Una antigua idea que sigue
investigándose en la actualidad, es utilizar el factor de
crecimiento similar a la insulina (IGF-1) para estimular el
crecimiento de los miocardiocitos. De hecho, los bajos niveles
circulantes en sangre de IGF-1 podrían ser
factor de riesgo cardiovascular [véase
la revisión de Kaplan et al., Cardiol Rev 13:35-9
(2005)].
– La reprogramación de fibroblastos. Aunque
intuitivamente concebimos el corazón básicamente como
un músculo, lo cierto es que los miocardiocitos están
numéricamente en minoría. En una sección
histológica del corazón adulto los miocardiocitos
sólo constituyen alrededor de una quinta parte de las
células presentes. Los fibroblastos, las células
productoras del tejido conectivo, son mucho más abundantes y
por supuesto mucho más capaces de proliferar. De hecho, los
fibroblastos constituyen el tejido cicatricial después de un
infarto. La cuestión, de momento sólo en el ámbito
de la especulación científica, es ¿podríamos
«reprogramar» los fibroblastos cardiacos para su
transdiferenciación en miocardiocitos? No es una
cuestión fácil, pero sí que es cierto que los
fibroblastos tienen capacidad de transdiferenciación a
miofibroblastos, células que poseen características y
expresan marcadores de músculo liso. Hay que considerar
además que fibroblastos cardiacos y miocardiocitos son
derivados mesodérmicos que proceden de un linaje embrionario
común. Esto ilustra la importancia de conocer bien los
mecanismos embrionarios de diferenciación celular para
poder aplicar estos
conocimientos a cuestiones concretas de interés clínico. Y
constituye un buen ejemplo, otro más, de la convergencia
entre la Biología del Desarrollo y la Medicina.
13.
TEJIDO MUSCULAR LISO VASCULAR: POBLACIÓN Y COMUNICACIÓN
CELULAR.
El músculo liso carece del patrón de estriaciones
característico de los músculos esquelético
y cardíaco, pero como éstos puede desarrollar
tensión activa por la interacción de las
proteínas actina y
miosina, mediante un proceso que consume ATP.
Las células musculares lisas tienen forma de huso. El
diámetro máximo varía entre 5 y 10 mm, y la
longitud entre 20 y 200 mm. En su superficie presentan
cavéolas, invaginaciones de la membrana plasmática o
sarcolema. Las células están rodeadas de una matriz extracelular reticular
que une las células en haces que forman bandas, y contribuye
a determinar las propiedades elásticas del Músculo Liso
Vascular.
La población celular que presentan son:
1) Población celular de Fibras
Musculares Lisas Vasculares.
2) Población celular de
Miofiibroblastos.
Las células del Músculo Liso Vascular se conectan
mediante dos tipos de unión intercelular:
1) Las uniones adherentes o uniones estrechas entre las
fibras musculares lisas vasculares proporcionan anclajes
mecánicos, lo que evita que las células se separen
durante la contracción, así como también permite
la transmisión de la tensión pasiva y activa que
desarrollan. Estas uniones adherentes están formadas por
desmosomas y adhesiones focales, estas últimas en
mayoría, las cuales están formadas por proteínas
transmembranas integrinas que interaccionan con
proteínas reguladoras como cinasa o tirosincinasa de la
adhesión focal.
2) Las uniones comunicantes son estructuras formadas
por conexones que constituyen canales o conductos relativamente
amplios que establecen una continuidad entre el citoplasma de las
células adyacentes, lo cual permite el paso de sustancias
con una masa molecular de hasta 1000 Da y la propagación de
señales eléctricas
entre las células.
BIBLIOGRAFÍA:
- ROSS, Michel; PAWLINA, Wojciech. Histología, Texto y
Atlas color con Biología Celular y Molecular, 5ª
edición, Buenos Aires: Médica Panamericana,
2007. - D. W. FAWCETT. Tratado de
histologia 12° edicion, 1995 editorial McGraw hill
Interamericana. - http://cursos.juanncorpas.edu.co/file.php/58/TEJIDO_MUSCULAR/TEJIDOMUSCULAR.pdf
14. SARCÓMERA
LISA.- CONTRACCIÓN VASCULAR.
Sarcómera Lisa
Los componentes del aparato contráctil de las
células musculares lisas son:
Filamentos finos:
Están adheridos a densidades citoplasmáticas o
"cuerpos densos" que se ven entre los filamentos. Estos
filamentos contienen actina, la isoforma muscular lisa de
la tropomisina y dos proteínas con acción dependiente de
Ca2+ y la caldesmona y la calponina. La
actina participa en la interacción generadora de fuerza con las moléculas
de miosina II. La tropomiosina muscular lisa no tiene troponina
asociada y su posición sobre filamentos de actina está
regulada por la fosforilación de las cabezas de miosina de
caldesmona (120 a 150 KDa) y la calponina (34 KDa) son
proteínas fijadoras de actina que bloquean el sitio de
unión para la miosina.
Filamentos grasos:
Contienen miosina II. Esta miosina también
está compuesto por 2 cadenas polipeptídicas pesadas y 4
cadenas ligeras, pero en lugar de adquirir una organización bipolar las
moléculas de miosina, están orientadas en una dirección en un lado del
filamento y en la dirección opuesta en el otro lado. Este
"filamento de miosina polar lateral" tampoco tiene una
"región desnuda" central, sino que exhibe extremos
desnudos aguzados asimétricos.
Otras proteínas asociadas al aparato contráctil son
las cinasa de las cadenas ligeras de la miosina (MLCK) que inicia
el ciclo de contracción del músculo liso; la a-actinina
que forma el componente estructural de los cuerpos densos y la
CALMOLUDINA, proteína fijadora de Ca2+ que
forma el "Complejo Ca2+-calmodulina" para activar
la MLCK.
Contracción:
Las células musculares lisas carecen de un sistema T
Un aspecto característico de las células musculares
lisas es la presencia de gran cantidad de invaginaciones de la
membrana celular que parecen cavéolas. Debajo de la membrana
plasmática y con frecuencia en las cercanías de las
pocas cisternas del REL hay vesículas citoplasmáticas.
Se cree que las invaginadones de la membrana celular y las
vesículas subyacentes junto con el REL funcionan de una
manera análoga a la del sistema T del músculo
esquelético para entre Ca2+ al citoplasma.
Las concentraciones intracelulares de Ca2+ son muy
importantes para la regulación de la contracción del
músculo liso.
El aumento de la concentración intracelular de
Ca2+ en la célula muscular lisa se
logra mediante la despolarización de la membrana celular con
la activación ulterior de proteínas sensoras de voltaje
o mediante la activación directa de canales con compuerta
para la liberación de Ca2+ en el REL por una
molécula según mensajero, en general IP3-.
El receptor de IP3- está ubicado en la membrana
del REL y tiene propiedades semejantes a las de los canales con
compuerta para la liberación de Ca2+ La
cantidad de Ca2+ que entra en la célula
después de la activación de la proteína sensora de
voltaje suele ser insuficiente para iniciar la contracción
muscular lisa y debe suplementarse con la liberación de
Ca2+ desde el REL. Luego el Ca2+ se
une a la calmodulina, que activa la fosforilación de la
cinasa de las cadenas ligeras de la miosina para iniciar la
contracción. Después del comienzo del ciclo de la
contracción el Ca2+ es extraído del
sarcoplasma por bombas de calcio dependientes de
ATP y se vuelve a secuestrar en el REL o se envía al medio
extracelular.
15. CORAZON.
MORFOLOGÍA CARDÍACA.
CAVIDADES CARDÍACAS:
El corazón se divide en cuatro cavidades, dos superiores
o aurículas (o atrios) y dos inferiores o ventrículos.
Las aurículas reciben la sangre del sistema venoso, pasan a
los ventrículos y desde ahí salen a la circulación
arterial.
Los ventrículos tienen la forma de una cavidad conoidea,
cuyo vértice se dirige hacia la izquierda, abajo y adelante,
hacia la punta del corazón. La base, en cada uno de los dos
ventrículos, presenta dos orificios; el orificio
auriculoventricular u orificio venoso, y el otro, el orificio
arterial.
En las parades, la superficie interior de los ventriculos,
presenta todo un sistema de eminencias y prolongaciones de la
sustancia muscular, las cuales, entrecruzandose en todos
sentidos, dan a la pared ventricular un aspecto reticulado o
alveolar. Estas eminencias son las columnas carnosas del
corazón; estas son abundantes en la punta. (TESTUD-LATERJET,
1978)
Característica | Ventrículo Izquierdo | Ventrículo Derecho |
Dirección | Paralelo al eje medio | Inclinado sobre el eje medio |
Forma | Conoidea | Prismática triangular |
Espesor | Más gruesa(15 mm) | Menos gruesa(5 mm) |
Las aurículas se hallan encima de los ventrículos,
tienen paredes delgadas y ausencia de columnas carnosas de primer
orden. Presentan dos tipos de orificios; un orificio auriculo
ventricular y otro orificio venoso (desembocadura de conductos
venosos).
La forma de las aurículas son muy irregulares, por ello,
difícilmente comparables. No obstante se les puede
considerar como cuboideas y estudiarlas por sus seis caras o
paredes. (TESTUD-LATERJET, 1978)
La aurícula derecha y el ventrículo derecho forman
lo que clásicamente se denomina el corazón derecho.
Recibe la sangre que proviene de todo el cuerpo, que desemboca en
la aurícula derecha a través de las venas cavas
superior e inferior. Esta sangre, baja en oxígeno, llega al
ventrículo derecho, desde donde es enviada a la
circulación pulmonar por la arteria pulmonar. Dado que la
resistencia de la
circulación pulmonar es menor que la sistémica, la
fuerza que el ventrículo debe realizar es menor, razón
por la cual su tamaño es considerablemente menor al del
ventrículo izquierdo.
La aurícula izquierda y el ventrículo izquierdo
forman el llamado corazón izquierdo. Recibe la sangre de la
circulación pulmonar, que desemboca a través de las
cuatro venas pulmonares a la porción superior de la
aurícula izquierda. Esta sangre está oxigenada y
proviene de los pulmones. El ventrículo izquierdo la
envía por la arteria aorta para distribuirla por todo el
organismo.
El tejido que separa el corazón derecho del izquierdo se
denomina septo o tabique. Funcionalmente, se divide en dos partes
no separadas: la superior o tabique interauricular, y la
inferior o tabique interventricular. Este último es
especialmente importante, ya que por él discurre el
fascículo de His, que permite llevar el impulso a las partes
más bajas del corazón.
VÁLVULAS CARDÍACAS:
Las válvulas de corazón o
válvulas cardíacas son tejidos del tamaño de una
moneda grande, que se encuentran en los conductos de salida de
las cuatro cavidades del corazón donde cumplen la finalidad
de dejar pasar la sangre en la dirección correcta, evitando
que ésta fluya hacia atrás. Con las diferentes
contracciones del corazón, se contraen también en una
secuencia determinada las cuatro cavidades, bombeando la sangre
en una dirección. Sin las válvulas, la sangre
volvería a la cavidad después de la contracción,
con lo cual el corazón no cumpliría su misión.
Las cuatro válvulas del corazón son las
siguientes:
Válvula tricúspide: Esta está ubicada en
la unión de la aurícula derecha con el ventrículo
derecho. La válvula tricúspide evita el reflujo de
sangre hacia la aurícula derecha, ya que se cierra por
completo durante la contracción del ventrículo derecho.
Esta formada por tres valvas (membranas).
Válvula bicúspide o mitral: Se encuentra
ubicada entre el paso de la aurícula izquierda y el
ventrículo izquierdo. Impide que la sangre retorne del
ventrículo izquierdo a la aurícula izquierda. Está
formada por dos valvas.
Cada válvula auriculoventricular tiene valvas o
cúspides, cuyas bases se insertan en el anillo fibroso que
rodea el orificio. Las caras auriculares de estas valvas son
lisas, en tanto que las caras ventriculares son rugosas debido a
la inserción de las cuerdas tendinosas. Los bordes libres de
las valvas con frecuencia tienen pequeños engrosamientos
nodulares. Las fibras musculares de la aurícula y una red capilar pueden aparecer en las bases
de las valvas, pero se prolongan en su interior algunos
milímetros; el resto de las valvas se componen de tejido
conectivo denso, a vascular, recubiertas en cada cara por el
endocardio.
Válvula sigmoidea pulmonar: Impide que la sangre
retorne del conducto pulmonar al ventrículo derecho.
Está formada por tres membranas.
Válvula sigmoidea aórtica: Impide que la
sangre retorne desde la aorta al ventrículo izquierdo.
Está formada por tres membranas.
Cada válvula sigmoidea tienes tres valvas, constituidas
por tejido fibroso a vascular, el borde libre de cada valva tiene
un pequeño engrosamiento central de tejido fibroso, el
Nódulo, en la arteria pulmonar es el nódulo de Arancio
y para la arteria aorta, el nódulo de Morgagni. A cada lado
del nódulo se extiende un área estrecha delgada, en
forma de media luna, denominada lúnula, que carece de tejido
fibroso, los espacios entre las valvas y las paredes de los vasos
"distalmente" son los senos aórticos y pulmonares,
respectivamente.
16. HISTOGéNESIS
DEL CORAZÓN. Desarrollado por Cintya Karol Rodríguez
Martinez.
El primordio del corazón se observa por primera vez a los
18 días. En el área cardiógena, las células
mesenquimales esplácnicas ventrales respecto del celoma
pericárdico se agregan y se unen entre ellas para formar dos
primordios cardíacos, los cordones
angioblásticos. Estos cordones de células
mesenquimatosas, pronto se canalizan y dan lugar a dos
tubos cardiacos endocárdicos de pared delgada;
localizados en el piso de la futura cavidad pericárdica,
estos tubos pronto se fusionan para formar un tubo
cardiaco único.
Conforme se fusionan los tubos cardiacos, se forma una capa
externa del corazón embrionario, el miocardio
primitivo, a partir del mesodermo esplácnico que
circunda al celoma pericárdico. En esta fase, el
corazón está compuesto por un tubo endotelial delgado,
separado de un tubo muscular grueso, el miocardio primitivo, por
un tejido conjuntivo gelatinoso, la gelatina cardiaca. El
tubo endotelial se convierte en el revestimiento endotelial
interno del corazón o endocardio, mientras que el
miocardio primitivo constituye la pared muscular del corazón
o miocardio. El pericardio vascular o epicardio
deriva de células mesoteliales que surgen de la superficie
externa del seno venoso y se dispersan por el miocardio
A medida que tienen lugar el plegamiento de la región de
la cabeza, el corazón y la cavidad pericárdica se
sitúan en posición ventral respecto al intestino
anterior y caudal en relación a la membrana
bucofaríngea. Al mismo tiempo, el corazón tubular se
alarga y desarrolla dilataciones y constricciones alternas, las
cuales son:
1) Seno venoso, región
caudal del corazón primitivo que recibe toda la sangre que
regresa del corazón a partir de las venas cardinales
comunes, venas vitelinas y venas umbilicales
2) Aurícula
3) Ventrículo
4) Bulbo cardiaco
5) Tronco arterioso.
El tronco arterioso se continúa con el saco
aórtico en la zona craneal, del cual surgen los arcos
aórticos. El seno venoso recibe la vena
umbilical, vitelina y cardinal común. Los extremos arterial
y venoso del corazón se fijan a los arcos faríngeos y
septum transversum, respectivamente.

Puesto que el bulbo cardiaco y el ventrículo crecen con
mayor rapidez, el corazón se pliega sobre sí mismo y
forma un asa bulboventricular en forma de "U". A medida
que el corazón primitivo se dobla, la aurícula y el
seno venoso adquieren una porción dorsal respecto al tronco
arterioso, bulbo cardíaco y ventrículo.
En este momento, el seno venoso ha desarrollado expansiones
laterales, las astas derechas e izquierda del seno
venoso.
Conforme se alarga y dobla, el corazón se invagina de
modo gradual hacia la cavidad pericárdica.
Inicialmente, el corazón está suspendido de la pared
dorsal por un mesenterio, el mesocardio dorsal, pero la
parte central de este mesenterio degenera pronto, formando una
comunicación, el seno
pericárdico transversal, entre los lados derecho
e izquierdo de la cavidad pericárdica. En esta etapa, el
corazón se halla unido solamente por sus extremos craneal y
caudal.
Durante la cuarta y quinta semana, el corazón primitivo
se divide en el órgano humano típico de cuatro
cámaras.

Destino del seno venoso y formación de la
aurícula derecha adulta: El seno venoso al
principio es una cámara separada del corazón primitivo
que se abre hacia la aurícula derecha. Mientras
continúa el desarrollo del corazón, el cuerno
izquierdo del seno venoso se trasforma en el seno coronario,
y su cuerno derecho se incorpora en la pared del atrio o
aurícula derecha, donde forma la porción suave de la
pared atrial derecha adulta. La mitad derecha del atrio o
aurícula primitiva, persiste como orejuela derecha o
apéndice del atrio.
División del conducto
auriculoventricular: Se llevan a cabo
proliferaciones localizadas de mesénquima, llamadas
almohadillas o cojinetes endocárdicos, en la
región auriculoventricular del corazón. Estas
almohadillas crecen una hacia la otra y se fusionan y dividen el
conducto auriculoventricular en dos, derecho e izquierdo
(AV).
División del atrio o aurícula
primitiva: Una partición membranosa con
apariencia de media luna conocida como septum primum, se
desarrolla de la pared dorsal del atrio primitivo, este septum
primum se fusiona con las almohadillas endocárdicos.
Antes de que el septum primum se fusione con estas almohadillas,
hay una comunicación entre las mitades izquierda y
derecha del atrio primitivo a través del ostium primum o
foramen primum.
Mientras que el septum primum se fusiona con las almohadillas
endocárdicas, y cierra el foramen primum, la parte superior
del septum se rompe y crea otra apertura llamada foramen
secundum. A medida que se desarrolla este agujero, otro
pliegue membranoso en forma de media luna, llamado septum
secundum se forma en el atrio hacia la derecha del septum
primum. Hay también una abertura entre el borde libre del
septum secundum y la pared dorsal del atrio. Se le denomina
agujero oval. Hacia este estadio, los restos del septum primum
han formado una válvula con apariencia de colgajo sobre el
agujero oval.
Formación de los
ventrículos: El ventrículo primitivo, da
lugar a la mayor parte del ventrículo izquierdo, mientras el
bulbo cardiaco forma casi todo el ventrículo derecho. El
tabique interventricular se inicia como una puente entre el piso
del ventrículo primitivo, y crece en forma lenta hacia
las almohadillas endocárdicas. Hacia el final de la
séptima semana, los futuros ventrículos izquierdo y
derecho se comunican a través de un gran agujero
interventricular. El cierre de este último da como
resultado la formación de la parte membranosa del tabique
interventricular. Esta se deriva de la fusión de tejido de
las almohadillas endocárdicas y los puentes bulbares.
Partición de los bulbos cardiacos y troncos
arteriosos: La división de estas partes del
corazón primitivo, desarrolla y fusiona las crestas
troncales y crestas bulbares. Los puentes mesenquimatosos,
forman un tabique aorticopulmonar que divide el tronco
arterioso y el bulbo cardiaco en aorta ascendente y tronco
pulmonar.
Desarrollo del sistema de conducción del
corazón: El nódulo sinoauricular, se forma
al principio en la pared del seno venoso, cerca de su apertura
hacia la aurícula derecha. El modulo sinoauricular se
incorpora más tarde en la aurícula derecha con el
cuerno derecho del seno venoso. El nódulo y el haz
auriculoventricular se derivan de células en las paredes del
seno venoso y el conducto auriculoventricular.ç
BIBLIOGRAFÍA:
- MOORE PERSAUD, " EMBRIOLOGÍA CLINICA", 7º
edición. Editorial GEA Consultoría Editorial,
S.L.L. Madrid España 2004
17. HISTOLOGÍA DE LAS PAREDES CARDIACAS:
AURÍCULAS VENTRÍCULOS.
(TABIQUES)
La organización estructural de la pared del
corazón es continua en las aurículas como en los
ventrículos; pero con algunas diferencias debido al trabajo que realizan.
Las capas de la cual se compone de tres capas las cuales
podemos mencionar:
Epicardio
Endocardio
Miocardio
EL PERICARDIO:
Está compuesto por dos hojas el epicardio parietal y el
visceral:
Pericardio parietal: se compone de tejido
fibrocolagenoso compacto y tejido elástico y que esta
tapizado en su interior por una capa de células
mesoteliales planas,
El pericardio visceral: también es llamado
epicardio, conforme ala cubierta externa del corazón y tiene
una capa externa de células mesoteliales planas. Estas
células descansan sobre un estroma de tejido fibrocolagenoso
de sostén que contiene fibras elásticas, así como
grandes vasos, también posee una serie de ganglios.
MIOCARDIO:
El miocardio es una capa de músculo estriado
especializado.
La cantidad de miocardio y el diámetro de las fibras
musculares en las fibras del corazón varia de acuerdo con
el trabajo al que se ve
sometida la cámara.
Aurícula derecha e izquierda:
Dado que empujan la sangre y la introducen en los
ventrículos vacíos, venciendo una mínima
resistencia durante su diástole .por ello su pared es
delgada y esta compuesta por células delgadas.
La superficie externa del miocardio que se encuentra por
debajo del pericardio es lisa, pero la superficie
interna por debajo del endocardio esta llena de
trabeculaciones que están más marcadas en los
ventrículos, además las trabeculaciones están
cubiertas de endocardio liso.
Las fibras atriales miocárdicas contienen unos
pequeños gránulos neuroendocrinos; una
característica es que estos gránulos se ubican en mayor
cantidad en la aurícula derecha. Estos gránulos
segregan la hormona natriurética auricular cuando se elongan
excesivamente las fibras auriculares.
El ventrículo derecho:
Debido a que impulsa la sangre a través de la
válvula pulmonar y los pulmones hasta que entra en la
aurícula izquierda. Por ello, posee una capa muscular
moderadamente gruesa compuesta por fibras de diámetro
intermedio entre las células musculares auriculares y
ventriculares.
Ventrículo izquierdo:
Bombea la sangre a un sistema arterial de gran presión y por tanto su pared
es mas gruesa y sus fibras musculares so las de mayor
diámetro.
En ambos ventrículos se levantan uno s cuantos
montículos de músculo cardiaco (músculos
papilares) que se protruyen en la luz de los ventrículos y que
apuntan hacia las válvulas auriculoventriculares. Los
músculos papilares son el lugar de inserción de las
cuerdas tendinosas, cordones tendinosos estrechos que
atan a las hojas de la válvula
auriculoventricular a la pared del ventrículo
por debajo de las mismas.
EL ENDOCARDIO:
El endocardio es variable en cuanto a grosor, siendo
más grueso en las aurículas y mas delgado
en los ventrículos particularmente en el ventrículo
izquierdo.
Esta estructura del corazón
esta compuesta por tres capas:
La capa externa, también llamada
subendocárdica, esta compuesta por fibras de colágeno
dispuestas irregularmente. Esta capa también puede
poseer algunas fibras de purkinje, que forman parte del sistema
de conducción.
La capa media, es la capa endocárdica más
gruesa y esta compuesta por unas fibras de colágeno
organizadas regularmente que contienes fibras elásticas en
luna configuración paralela a la parte mas profunda de
la capa, también posee células del músculo
liso,
La capa interna, esta compuesta por células
endoteliales planas y acompañada de un tejido conjuntivo
subendotelial.
TABIQUES DEL CORAZON:
Tabique interventricular, es la pared que separa el
ventrículo derecho de ventrículo izquierdo, además
contiene:
Ø Músculo cardiaco en su porción
membranosa.
Ø Tapizado por endocardio.
Tabique interauricular, mucho más delgada que la
anterior; compuesta por:
Ø Tejido fibroso.
Ø Una capa central de músculo cardiaco.
Ø Un revestimiento d endocardio en la superficie de
contacto con la cavidad.
BIBLIOGRAFÍA:
- Alan Stevens; James Steven Lowe. 2006. Histología
Humana. Tercera edición. Editorial Elsevier.
Madrid-España.
- Junqueira Luiz Carlos; José Carneiro. 2000.
Histología Básica. Quinta edición. Editorial
Masson, Barcelona-España
·
Bloom William; Fawcett Wayne Don.1995. Tratado de
Histología. Duodécima Edición. Editorial
McGraw-Hill Interamericana. Madrid-España
18. HISTOLOGIA DEL APARATO MITRAL Y
DE LAS VÁLVULAS CARDIACAS.
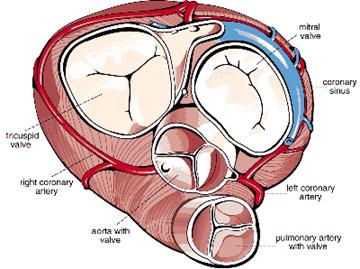
La válvula de entrada en el ventrículo derecho esta
formada por tres hojuelas y se denomina tricúspide. La de
ingreso del ventriculo izquierdo esta formada por dos hojuelas;
por lo tanto, se llama válvula bicúspide (o mitral).
Las hojuelas de las dos válvulas auriculoventriculares
tienen una estructura histológica similar. (Ham, 1982)
VALVULAS CARDIACAS
Las valvulas cardiacas, estan
formandas por valvas u hojuelas, las cuales poseen una capa media
de soporte formada por tejido conectivo colageno denso recubierta
en ambos lados por endocardio. Esta capa media se continua con el
tejido conectivo denso de los anillos fibrosos que rodean los
orificios. En la cara ventricular de la capa de tejido colageno
hay un gran numero de fibras elasticas , tambien las hay en un
número limitado debajo del endotelio en la capa auricular de
la hojuela.
Se han descrito a este nivel fibras musculares lisas y
se les ha atribuído acción de esfínter.
Desde los músculos papilares se extienden cuerdas
tendinosas de tejido conectivo colágeno denso
recubiertas de endocardio delgado que van a parar a la cara
ventricular de la capa media que soporta cada valva.
(Ham, 1982)
La valvula de salida del ventriculo derecho reciben el nombre
de valvula semilunar pulmonar, y la del ventriculo izquierdo es
la valvula semilunar aortica; ambas estan formadas por tres
hojuelas.
Las hojuelas de las valvulas semilunares son mas delgadas que
las de las valvulas auriculoventriculares. Sin embargo tienen
igual estructura general; estan formadas por pliegues de
endocardio reforzadas por una capa media de tejido conectivo
denso; los pliegues de endocardio en su base se continuan con el
esqueleto del corazón .
No tienen cuerdas tendinosas.
(Ham, 1982)
Estableciendo una estructura general de las válvulas
cardiacas, cada una de estas está compuesta por tres
capas:
v Fibrosa: forma el centro de cada valva y
contiene extensiones fibrosas del tejido conjuntivo denso
irregular de los anillos fibrosos.
v Esponjosa: formado por tejido conjuntivo laxo
ubicado en el lado auricular de la válvula (válvulas
tricúspide y mitral) y en el lado del vaso sanguíneo
(válvulas aórtica y pulmonar). Esta capa actúa
como amortiguador porque reduce las vibraciones asociadas con el
cierre de la válvula, también confiere elasticidad y plasticidad a la
válvula.
v Ventricular: Al lado de la superficie
ventricular de cada valva y tiene revestimiento endotelial.
Contiene tejido conjuntivo denso con muchas capas de fibras
elásticas. Esta es continua con las cuerdas tendinosas,
finos cordones fibrosos revestidos con endotelio.

APARATO MITRAL
El aparato mitral esta formado por el anillo fibroso, las
cuerdas tendinosas, los musculos papilares, las
valvas, y la pared ventricular izquierda asociada.
(Kumar-Abbas-Fausto, 2005)
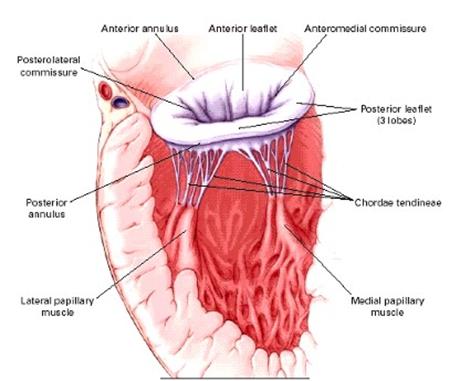
Estructura histológica de la valva mitral:
Desde el punto de vista histológico, podemos dividir a la
válvula mitral normal en cuatro capas, que desde el atrio al
ventrículo son:
- Superficie atrialis (endocardio, continuación del
atrial). - Pars spongiosa, que se trata de una colección de
escasas fibras y haces de colágeno, y algunas fibras
elásticas embebidas en una sustancia fundamental de
mucopolisacáridos. - Pars fibrosa, constituye el esqueleto valvular, y está
constituido por una densa capa de fibras de colágeno, que
se continúan en apical con el anillo mitral y en distal
con las cuerdas tendinosas. - Superficie ventricularis (endocardio que se continua con el
revestimiento ventricular)

BIBLIOGRAFIA:
- Arthur W. Ham: Tratado de Histología. 7º
edición. Editorial Interamericana. Mexico, D.F , 1982 - Autor Vinay Kumar, Stanley L. Robbins, Abul K. Abbas,
Nelson Fausto: Patología estructural y funcional. 7º
edición. Editorial Elsevier. 2005 - www.fcv.unlp.edu.ar/sitios-catedras/5/material/Peq-Practico-2005-Charla_ivm.doc
19. SISTEMA DE CONDUCCION
CARDIACA – HISTOFISIOLOGÍA.
Este sistema se compone de células musculares
cardiacas y fibras de conducción (no se trata de tejido
nervioso sino que son el resultado de impulsos generados en el
propio corazón), que están especializadas en el inicio
de los impulsos y en su conducción rápida a través
del corazón.
Este sistema inicia los latidos normales del corazón y
coordina las contracciones de las cuatro cavidades cardiacas. Las
dos aurículas se contraen al mismo tiempo, al igual que los
ventrículos, pero la contracción auricular ocurre
antes. El sistema de conducción otorga al corazón este
automatismo y ritmicidad en los latidos. Para que el corazón
bombee eficazmente la sangre y puedan actuar sincrónicamente
las circulaciones sistémica y pulmonar, es necesario
coordinar todos los acontecimientos del ciclo cardiaco.
El nódulo sinusal (nódulo SA) Esta
agrupación de fibras especializadas de musculo cardiaco
(tejido nodal) en la pared de la aurícula derecha inicia los
impulsos para la contracción del corazón. El
nódulo sinusal es el marcapasos natural del
corazón. El nódulo SA está situado en el
extremo superior de la cresta terminal, en la unión de la
cara anteromedial de la vena cava superior con la orejuela
derecha. Se puede localizar su posición, siguiendo la
arteria para el nódulo SA, rama de la arteria coronaria
derecha (a veces de la izquierda). El nódulo SA recibe
inervación de las dos divisiones del SNA (sistema nervioso
autónomo). El nódulo emite impulsos con una frecuencia
aproximada de 70 impulsos por minuto en la mayoría de
personas. La velocidad con la que se
producen los impulsos en el nódulo puede variar por
estimulación nerviosa: la estimulación simpática
acelera la frecuencia y la estimulación parasimpática,
la frena.
El nódulo auriculoventricular (nódulo AV)
Esta agrupación de tejido nodal, mucho más
pequeña, se compone de células especializadas de
musculo cardiaco y está situada en el tabique
interauricular, en la cara ventricular del orificio del seno
coronario. Los impulsos de las fibras de musculo cardiaco de las
dos aurículas convergen en el nódulo AV, que los
distribuye por los ventrículos a través del
fascículo auriculoventricular. El nódulo AV conduce los
impulsos más lentamente: la estimulación simpática
acelera la conducción y la parasimpática, la frena.
El fascículo auriculoventricular (fascículo
AV) Llamado comúnmente Haz de His, el Haz de His,
sus ramas y las fibras de Purkinje están compuestos por
células musculares cardiacas modificadas que se han
especializado en la conducción de impulsos, ésta
agrupación de fibras especializadas del musculo de
conducción, que se denomina a menudo fibras de Purkinje se
originan en el nódulo AV y discurre por la porción
membranosa del tabique interventricular. El fascículo AV se
divide en la unión entre las porciones muscular y membranosa
del tabique interventricular en sus ramas derechas e izquierda.
Cada rama camina en la profundidad del endocardio, por las
paredes de los ventrículos. La rama derecha del
fascículo AV inerva el musculo del tabique interventricular,
el musculo papilar anterior y la pared del ventrículo
derecho. La rama izquierda del fascículo AV se distribuye
por el tabique interventricular, los músculos
papilares y la pared del ventrículo izquierdo.
Los nódulos y el has de Hiz y sus ramas contienen
células musculares cardiacas modificadas de un tamaño
menor que las normales. En cambio, las de Purkinje son
fibras musculares cardiacas modificadas más grandes que las
normales.

BIBLIOGRAFÍA:
- Keith L. Moore, "Anatomía con orientación
clínica", 3º Edición. Editorial, Editorial
medica Panamericana. 1995
20. BREVE REFERENCIA
HISTOLÓGICA SOBRE ARTERIA AORTA Y VENAS CAVAS.
La arteria Aorta posee, como todas las arterias y venas,
las tres capas o túnicas (íntima, media y adventicia),
pero se la clasifica como una arteria elástica y tiene
ciertas particularidades:
a) La túnica íntima de
la arteria Aorta (elástica) se compone de un endotelio,
tejido conjuntivo subendotelial y una membrana elástica
interna no conspicua:
La túnica íntima de la arteria Aorta es
relativamente gruesa y está compuesta por:
· Endotelio
de revestimiento: Con su lámina basal. Las células
son típicamente planas y alargadas y sus ejes mayores
están orientados paralelos a la dirección del flujo
sanguínea en la arteria. Para formar la lámina
epitelial las células están unidas por zonulae
occludentes y uniones de hendidura (nexos). Las células
endoteliales poseen en su citoplasma inclusiones bastoniformes
llamadas cuerpos de Weibel-Palade. Estas inclusiones endoteliales
específicas son estructuras electrodensas que contienen el
factor de Von Willebrand (también conocido como factor VIII
de la coagulación). Varios estudios indican que la mayor
parte del factor de von Willebrand es sintetizado por la
célula endotelial arterial y secretado hacia la sangre. Los
anticuerpos contra el factor Von Willebrand suelen usarse como
marcadores inmunohistoquímicos para la identificación
de los tumores derivados del endotelio.
· Capa
subendotelial: De tejido conjuntivo, que en las arterias
elásticas más grandes contiene tanto fibras
colágenas como fibras elásticas. El tipo celular
principal en esta capa es la célula muscular lisa. ésta
es contráctil y secreta sustancia fundamental extracelular,
así como fibras colágenas y elásticas.
También puede haber macrófagos ocasionales.
· Membrana
(lámina) elástica interna: Que en las arterias
elásticas no se distingue con claridad porque es una de las
muchas láminas elásticas en la pared del vaso. Suele
identificarse sólo porque es la lámina elástica
más interna de la pared arterial.
b) La túnica media de las
arterias elásticas consiste en múltiples capas de
células musculares lisas separadas por láminas
elásticas:
La túnica media es la más grande de las tres capas
de las arterias elásticas y se compone de:
·
Elastina: En la forma de láminas fenestradas entre
las capas de células musculares. Estas láminas adoptan
una disposición concéntrica, las fenestraciones en las
láminas facilitan la difusión de sustancias dentro de
la pared arterial. La cantidad y el espesor de las láminas
están relacionados con la tensión arterial y la edad.
Al nacimiento, la aorta no tiene casi ninguna lámina
elástica, pero en el adulto hay entre 40 a 70 de estas
láminas. En las personas con hipertensión arterial
aumentan tanto la cantidad como el espesor de las láminas
elásticas.
·
Células musculares Lisas: Distribuidas en capas. Las
células musculares lisas describen una espiral de poca
pendiente en relación con el eje longitudinal del vaso, por
lo tanto, en los cortes transversales de la arteria aparecen con
una distribución circular. Estas
células son fusiformes y tienen un núcleo alargado.
Están rodeadas por una lámina (basal) externa excepto
en donde se unen por nexos (uniones de hendidura). En la
túnica media no hay fibroblastos. Las células
musculares sintetizan el colágeno, la elastina y las
demás moléculas de la matriz extracelular. Además,
en respuesta a factores de crecimiento (por ejemplo PDGF, FGF)
producidos por las células endoteliales, las células
musculares pueden proliferar y migrar hacia la túnica
íntima contigua. Esta característica es importante en
la reparación normal de la pared vascular, así como en
procesos patológicos similares a los que ocurren en la
arterosclerosis.
· Fibras
colágenas y sustancia amorfa (moléculas de
proteoglucanos): Que son sintetizadas y secretados por las
células musculares lisas.
c) La túnica adventicia en la
arteria elástica es una capa de tejido conjuntivo de
relativa delgadez:
En las arterias elásticas, la túnica adventicia
suele tener menos de la mitad del espesor de las túnica
media. Consiste en:
· Fibras
colágenas y fibras elásticas: Estas últimas en
la forma de una red fibrilar laxa (pero láminas) que
está menos organizada que la de la túnica media. Las
fibras colágenas contribuyen a impedir la distensión de
la pared arterial más allá de los límites fisiológicos
durante la sístole del ciclo cardíaco.
·
Fibroblastos y macrófagos: Principales células
de la túnica adventicia.
· Vasos
sanguíneos (vasa vasorum) y nervios (nervi vascularis).
Las ramas de los vasa vasorum penetran parcialmente la
túnica media y proveen sustancias nutritivas a la
porción externa de la pared vascular. La porción
interna de la pared recibe los nutrientes desde la luz del
vaso.
Las venas cavas poseen, como todas las arterias y
venas, las tres capas o túnicas (íntima, media y
adventicia), pero se las clasifica como una vena grande y tiene
ciertas particularidades:
En las venas cavas la túnica media es relativamente
delgada y la túnica adventicia es relativamente gruesa.
Las venas cavas tienen diámetro superior a 10 mm (venas
grandes).
a) La túnica íntima de
estas venas consiste en un revestimiento endotelial con su
lámina basal, una pequeña cantidad de tejido conjuntivo
subendotelial y algunas células musculares lisas. Con
frecuencia el límite entre la túnica íntima y la
túnica media no es claro y no siempre resulta fácil
decidir si las células musculares lisas cercanas al
endotelio pertenecen a la primera o a la segunda.
b) La túnica media es
relativamente delgada y contiene células musculares lisas de
disposición circunferencial, fibras colágenas y algunos
fibroblastos.
c) La túnica adventicia de las
venas grandes (venas cavas) es la capa más gruesa de la
pared vascular. Junto con las fibras colágenas, las fibras
elásticas y los fibroblastos habituales. La túnica
adventicia también contiene células musculares lisas de
disposición longitudinal.
20.
ARTERIAS CORONARIAS: DISTRIBUCIÓN ANATÓMICA E
HISTOFISIOLÓGICA.
Desarrollado por Diego Edilberto Morales
Fernández
A. Consideraciones generales
Las dos arterias coronarias principales son la coronaria
izquierda y la coronaria derecha. La arteria coronaria izquierda,
que se divide en las ramas: descendente anterior y circunfleja,
lleva la sangre a el ventrículo izquierdo del
corazón y al atrio izquierdo. La arteria coronaria derecha,
que se divide en la arteria descendente posterior derecha y la
arteria marginal aguda, irriga sangre al ventrículo derecho,
la aurícula derecha, el nódulo sinoauricular (grupo de células
localizado en la pared de la aurícula derecha que regula el
ritmo de los latidos del corazón), y nodo
auriculoventricular.
Las dos arterias coronarias principales tienen ramas
adicionales para llevar la sangre al músculo cardiaco. Estas
incluyen las siguientes:
· Arteria
circunfleja
Las arteria circunfleja sale de la arteria coronaria izquierda y
rodea el músculo cardiaco. Esta arteria lleva la sangre al
lado y a la parte trasera del corazón.
· Arteria
descendente anterior izquierda
La arteria descendente anterior izquierda sale de la arteria
coronaria izquierda y lleva sangre a la parte delantera de la
izquierda del corazón.
· Las
ramas más pequeñas de las arterias coronarias incluyen
las siguientes: marginal aguda, descendente posterior, marginal
oblicua, perforador septal y diagonales.
·
Distribución anatómica Las dos arterias
coronarias que irrigan al miocardio surgen de los senos de las
cúspides posteriores de la válvula aórtica en la
raíz de la aorta. Corrientes turbulentas mantienen las
valvas alejadas de los orificios arteriales, que permanecen
permeables durante todo el ciclo cardiaco. La arteria coronaria
derecha tiene mayor flujo en 50 % de los individuos, la izquierda
tiene mayor flujo en 20% y en el 30 % restante el flujo es igual
en ambas. La mayor parte de la sangre venosa regresa al
corazón por el seno coronario y las venas cardiacas
anteriores, las cuales drenan en la aurícula derecha.
Además, hay otros vasos que se vacía en forma directa
en las cámaras cardiacas. Estos incluyen los vasos
arteriosinusoidales, que son vasos sinusoidales similares a
capilares que conectan las arteriolas con las cámaras
cardiacas; las venas de Tebesio, que conectan los capilares con
las cámaras, y unos cuantos vasos arterioluminales que son
pequeñas arterias que drenan directamente en las
cámaras. Existen pocas anastomosis entre las arteriolas
coronarias y las arteriolas extracardiacas, sobretodo alrededor
de la abertura de las grandes venas. Las anastomosis entre las
arteriolas coronarias en los humanos sólo permite el paso de
partículas menores de 40 um de diámetro, pero hay
evidencia de que estos conductos aumentan de tamaño y
número en pacientes con cardiopatía isquémica.

B. Aspectos funcionales
Gradiente de presión y flujo den vasos
coronarios
El corazón es un músculo que, al igual que el
esquelético, comprime sus vasos sanguíneos cuando se
contrae. La presión dentro del ventrículo izquierdo es
un poco mayor que la presión aórtica durante la
sístole. Por consiguiente, sólo hay flujo en las
arterias que irrigan la porción subendocárdica
del ventrículo izquierdo durante la diástole, aunque la
fuerza se disipa lo suficiente en las porciones más
2superficiales del miocardio ventricular izquierdo
para permitir cierto flujo en esta región durante todo el
ciclo cardiaco. Ya que la diástole es más corta cuando
la frecuencia cardiaca es alta, el flujo coronario del
ventrículo izquierdo disminuye durante la taquicardia, Por
otro lado, la diferencia de presión entre la aorta y el
ventrículo derecho, y entre la aorta y las aurículas es
un poco mayor durante la sístole que en el transcurso de la
diástole. Como consecuencia, el flujo coronario en esas
partes del corazón no disminuye de manera apreciable durante
la sístole. Como no hay flujo de sangre durante la
sístole en la porción subendocárdica del
ventrículo izquierdo, esta región es proclive al
daño isquémico y es
el sitio más frecuente de infartos miocárdicos. El
flujo sanguíneo al ventrículo izquierdo disminuye en
pacientes con estenosis de la válvula aórtica porque en
este caso la presión del ventrículo izquierdo debe ser
mucho más alta que la aórtica para expulsar la sangre.
Por consiguiente, los vasos coronarios se comprimen mucho durante
la sístole. Los pacientes con este trastorno tienen
tendencia a presentar síntomas de isquemia miocárdica,
en parte por esta compresión y en parte porque el miocardio
requiere más O2 para expulsar la sangre por la
válvula aórtica estrecha. El flujo coronario
también disminuye cuando la presión diastólica
aórtica es baja. El incremento en la presión venosa en
padecimientos como la insuficiencia cardiaca congestiva disminuye
el flujo coronario porque reduce la presión de
perfusión coronaria efectiva.
Variaciones en el flujo coronario
En reposo, el corazón extrae 70 a 80% del O2
de cada unidad de sangre que le llega. El consumo de O2 puede
incrementarse mucho tan sólo con el aumento del flujo
sanguíneo. Por tanto, no es sorprendente que el flujo
sanguíneo aumente cuando el metabolismo del miocardio se
eleva. El calibre de los vasos coronarios y, por consiguiente, la
velocidad del flujo coronario están bajo la influencia no
sólo de los cambios en la presión en la aorta, sino
también de factores químicos y neurales. La
circulación coronaria tiene una autorregulación
considerable
Factores químicos
La estrecha relación del flujo sanguíneo coronario y
el consumo miocárdico de O2 indica que uno
o más de los productos del metabolismo
causan vasodilatación coronaria. Lo posibles factores que
ejercen este efecto incluyen la falta de O2 y el
aumento en las concentraciones locales de CO2,
H+, K+, lactato, prostaglandinas,
nucleótidos de adenina y adenosina. Es posible que
participen más de uno de estos metabolitos vasodilatadores.
La asfixia, la hipoxia y las inyecciones intracoronarias de
cianuro aumentan el flujo sanguíneo coronario 200 a 300% en
corazones desnervados y también en los normales, y el factor
común a estos tres estímulos es la hipoxia de las
fibras miocárdicas. Se produce un aumento similar en el
flujo en el área irrigada por una arteria coronaria si
ésta se ocluye y luego se libera. Esta hiperemia reactiva es
similar a la que se observa en la piel. Hay evidencia de que, el
corazón, este efecto se debe a la liberación de
adenosina. Parece que la adenosina además aminora la
lesión inducida por la reperfusión que ocurre cuando el
flujo sanguíneo se restablece.
Factores neurales:
Las arterias coronarias poseen una doble inervación,
simpática y parasimpática. E1 simpático, que en la
mayor parte del organismo es vasoconstrictor, ejerce sobre las
coronarias una acción vasodilatadora. El efecto
vasoconstrictor del parasimpático es, por el contrario, muy
discutido. Es importante recordar que tanto el vago como el
simpático modifican también la actividad cardíaca
y, por lo tanto, el consumo de O2 y la producción local de
metabolitos. Esto hace particularmente difícil evaluar la
participación que cabe a cada uno de estos factores en la
regulación de la irrigación cardíaca. Parece
probable que tanto la pO2 como la concentración
de catabolitos y la acción directa de los nervios,
actúen coordinadamente en dicha regulación.
Las arteriolas coronarias cuentan con receptores α
adrenérgicos que median la vasoconstricción y los
receptores β adrenérgicos que median la
vasodilatación. La actividad en los nervios
noradrenérgicos que llegan al corazón y las inyecciones
de noradrenalina causan vasodilatación coronaria. Sin
embargo, la noradrenalina aumenta la frecuencia cardiaca y la
fuerza de contracción, y la vasodilatación se
debe a la producción de metabolitos vasodilatadores en el
miocardio por el aumento en su actividad.
Cuando la presión arterial sistémica decae, el
efecto general del aumento reflejo en la descarga
noradrenérgica es el aumento del flujo sanguíneo
coronario por los cambios metabólicos en el miocardio en un
momento en que los vasos cutáneos, renales y
esplácnicos están constreñidos. De esta manera, se
conserva la circulación del corazón, al igual que la
del cerebro, cuando se compromete el
flujo sanguíneo a otros órganos.
22. VASOS INTRAMURALES:
HISTOLOGÍA Y FUNCIÓN EN SÍSTOLE Y EN
DIÁSTOLE.

"…Las arterias coronarias principales y sus ramas
epicárdicas normalmente sufren escasos cambios de calibre,
pero las pequeñas arteriolas intramiocárdicas tienen
una gran capacidad de dilatarse y cerrarse para regular el aporte
de sangre y de oxígeno en las diferentes situaciones
fisiológicas. Se cree que, debido a que los vasos quedan
parcialmente comprimidos por la contracción del miocardio
durante la sístole, la mayor parte del flujo se realiza
durante la diástole.
En los sujetos mayores las arterias coronarias son
especialmente vulnerables a la aterosclerosis. El depósito
subíntimo de células y lípidos conduce a la
formación de placas ateroscleróticas que reducen el
área de corte transversal de las arterias coronarias
epicárdicas. Cuando este proceso progresa hasta el punto de
que el flujo se hace insuficiente para garantizar a las
pequeñas ramas una respuesta al incremento en las demandas
de oxígeno el paciente puede experimentar una crisis de angina. El
estrechamiento aún mayor puede dar lugar a la isquemia de
una mayor zona del miocardio ventricular y a un infarto de
miocardio fatal…"
(Tratado de histología Bloom Fawcett. 12ª
edición 1996. Editorial mcgraw-hill
interamericana-pp449)
FUNCIÓN EN SÍSTOLE Y
DIÁSTOLE:
Las sístoles y las diástoles se suceden
alternativamente en forma rítmica y constituyen un ciclo
denominado revolución cardiaca, que se
cumple en 8/10 segundos
Ø Sístole:
Sístole es la fase activa (consumo de O2) del ciclo
cardíaco; se inicia con la contracción ventricular que
eleva la presión hasta cruzar la existente en la
aurícula izquierda, ahí inicia la fase de
contracción isovolumétrica; cuando cruza la
presión diastólica aórtica, se inicia la fase
expulsiva, se reduce el volumen intra-ventricular hasta
el fin de la fase expulsiva (volumen sistólico final),
después del cierre aórtico sobreviene la
relajación isovolumétrica; de tal manera de que la
sístole se compone esencialmente de 3 fases:
contracción isovolumétrica, fase expulsiva y
relajación isovolumétrica. (Raukin S, Lucke EJ, Gaynor
W, Harpole DH, Feneley MP, Nikolic S, et al: An Energetic
Analysis of Myocardial Performance In: Cardiac Mechanics and
Function in the Normal and Diseased Heart. (Eds. Hori M, Suga
H, Baan J, Yellin EL). Springer-Velay. 1989: 165-188.)

Ø Diastole:
Es la fase en la que el corazón se relaja., haciendo que
la sangre entre en las aurículas y se inicia de nuevo el
proceso.
Consiste en un adelgazamiento de la pared afectada con
engrosamiento de la pared sana, en el momento en que el
corazón se relaja. Se puede comprobar que si en vez de
ocluir la arteria, se la perfunde con una solución libre de
oxígeno, el fenómeno no se observa, demostrándose
que el disbalance interparietal observado es un fenómeno
mecánico consecuencia de la ausencia de un "esqueleto
hidráulico" en la pared afectada y no metabólico por la
anoxia tisular.
"…La estructura de las arterias elásticas permite
que sus paredes se distiendan durante la sístole
(contracción cardiaca), seguida del retroceso de las mismas
durante la diástole (relajación cardiaca), lo que ayuda
a conservar una presión arterial y un flujo sanguíneo
más constante. Las arterias musculares que se ramifican de
las arterias distribuyen sangre al cuerpo y están sujetas a
cambios constantes del diámetro que resultan de
vasoconstricción y vasodilatación. A fin de ayudar en
el ajuste para estos fenómenos, la túnica adventicia se
funde laxamente en el tejido conectivo circundante y por tanto
previene las restricción del vaso durante las contracciones
y expansiones para los cambios en la presión
arterial…."
Sístole

Diástole

"…El primero en ocuparse del movimiento de la sangre en las
coronarias fue, al parecer, Giovanni Battista Scaramucci. En su
libro de 168945 escribió
que, contrariamente a lo que ocurre en la circulación mayor,
en la cardiaca el flujo sanguíneo avanza esencialmente
durante la diástole, mientras que en la sístole, debido
a la compresión ejercida por las fibras miocárdicas
sobre los vasos, pueden llegar al miocardio sólo
pequeñas cantidades de sangre. En efecto, los vasos
intramurales se hallaban vacios durante la sístole. Lancisi
opinaba que la circulación coronaria se debe al impulso
sistólico y Rébatel demostró el sincronismo del
movimiento cardiaco con el coronario. Se cree actualmente
que el flujo sanguíneo por las coronarias es de tipo
continuo, aunque con variaciones i.e. con cierta reducción
durante la sístole. Debe tenerse presente que las regiones
endocárdicas y subendocardicas reciben sangre, con mecanismo
osmótico, de las cavidades ventriculares repletas de sangre
(diástole), mientras que las regiones epicárdicas y
subepicárdicas la reciben por los vasos coronarios tanto en
diástole como en sístole. El problema de la
función del flujo coronario ha llamado la atención de los
investigadores desde hace siglos…" "Scaramucci GB: De
motu cordis mechanicum theorema. Senigallia, 1689"
v Correlaciones Clínicas:
Lundbaek,11 en 1954, notó que en un mismo paciente
diabético se presentaban: retinopatía, nefropatía,
enfermedad cardíaca y enfermedad vascular periférica y
sugirió que estas lesiones vasculares podrían
constituir el síndrome de angiopatía diabética. El
mismo autor, en estudio postmortem encontró que los cambios
histológicos en los vasos intramurales pequeños del
corazón eran similares a los observados en otros tejidos de
pacientes con diabetes de larga duración y
propuso el término cardiopatía diabética para
describir la enfermedad cardíaca o disfunción
cardíaca en pacientes con diabetes mellitus, consideró
que ésta podía ser causada por una combinación de
enfermedad arterial de los grandes vasos coronarios,
neuropatía autonómica diabética y enfermedad de
las arterias y arteriolas coronarias pequeñas distales.
La existencia de miocardiopatía diabética fue
planteada por primera vez por Rubler y otros13 en 1972, y el
término fue introducido por Hamby y otros14 en 1974,
después de estudiar 73 pacientes con enfermedad
miocárdica primaria idiopática y constatar que de ellos
16 eran diabéticos, además comprobaron en el examen
postmortem de 3 pacientes diabéticos, ausencia de
aterosclerosis coronaria con lesiones no ateroscleróticas
presentes en los pequeños vasos intramurales. Los resultados
de los estudios de Framingham ofrecen evidencias posteriores que
apoyan la existencia de una enfermedad cardíaca
específica de la diabetes, en ellos se comprobó un
aumento de la frecuencia de fallo cardíaco congestivo en
diabéticos en ausencia de enfermedad arterial coronaria.
BIBLIOGRAFÍA:
- Tratado de histología Bloom Fawcett. 12ª
edición 1996. Editorial mcgraw-hill
interamericana-pp449
- Raukin S, Lucke EJ, Gaynor W, Harpole DH, Feneley MP,
Nikolic S, et al: An Energetic Analysis of Myocardial
Performance In: Cardiac Mechanics and Function in the Normal
and Diseased Heart. (Eds. Hori M, Suga H, Baan J, Yellin
EL). Springer-Velay. 1989: 165-188.
- "GARTNER Leslie. HISTOLOGÍA TEXTO Y ATLAS. 2ª
edición. Ed. Mc Graw-Hill Interamericana. 2002, pp250
- "Scaramucci GB: De motu cordis mechanicum theorema.
Senigallia, 1689"
23. DRENAJE LINFÁTICO E
INNERVACIÓN DEL CORAZÓN DRENAJE LINFÁTICO
CARDIACO.
Los capilares linfáticos drenan a los vasos en el
epicardio, donde siguen a las arterias coronarias y terminan en
los troncos colectores derecho e izquierdo. El tronco derecho se
dirige a los ganglios mediastinicos superiores (anteriores).El
izquierdo se dirige a un ganglio (el "ganglio caval") del grupo
traqueo bronquial superior, entre la aorta y la vena cava
superior.
INERVACIÓN CARDIACA
El corazón es inervado por fibras nerviosas
autónomas y por fibras sensitivas procedentes de los
nervios vagos y de los troncos simpáticos.
En las aurículas hay muchas células
ganglionares, particularmente cerca de los ganglios y las
venas. También se presentan algunas en los
ventrículos .Estas células nerviosas, siendo
parasimpático la mayor parte, se presentan tanto en el
epicardio como en el miocardio. Las terminaciones nerviosas
sensitivas complejas se encuentran en el corazón, sobre todo
en las aurículas cerca de la desembocadura de las venas y en
las paredes de las grandes venas.
FIBRAS AUTONOMAS: Las fibras simpáticas
preganglionres se originan del I al IV niveles
dorsales de la medula espinal..Hacen sinapsis en los ganglios
cervicales y dorsales y las fibras postganglionares que van
al corazón, son conducidos por ramas cardiacas de las
porciones cervical y dorsal del tronco simpático. Las
fibras parasimpáticas preganglionares en los nervios
vagos son conducidas por las ramas cardiacas cervical y
dorsal de los nervios vagos hacia las células
ganglionares en el corazón. Las fibras posganglionares de
ambos sistemas inervan el nódulo
de Keith y Flack (sinoauricular) y el de Aschoff y
Tawara (auriculoventricular) y los vasos coronarios.
Los nervios cardiacos son variables en su topografía y están
agrupados según su nivel de origen en la siguiente
forma:
- Nervios Cardiacos Cervicales (superior y medio): se
originan en el tronco simpático cervical, en los ganglios
o en ambos y generalmente se unen por ramas cardiacas
cervicales del vago. Después el conjunto de nervios
desciende por delante y por detrás del cayado
aórtico y entra al plexo cardiaco. - Nervios Cervicotoracicos(o cardiacos cervicales
inferiores): se originan en el ganglio cervicotoracico y el
asa subclavia, y generalmente se unen por medio de las ramas
cardiacas cervicotoracicos del vago. Después el conjunto
de nervios se dirige por delante o por detrás
del cayado aórtico hacia al plexo cardiaco. - Nervios Cardiacos Toracicos: se originan en el
IV o V ganglio simpáticos toracicos superiores y
junto con las ramas cardiacas torácicas del vago y los
nervios laringeos recurrentes izquierdos van directamente al
plexo cardiaco, especialmente a las paredes posteriores de las
aurículas.
Fibras Sensitivas: Las principales fibras de las
terminaciones sensitivas complejas en el corazón ascienden
por los nervios vagos. Las terminaciones libres se presentan en
el tejido conectivo del corazón y en la adventicia de los
vasos sanguíneos. Sus fibras principales se dirigen a las
porciones torácicas y cervicales inferiores de los troncos
simpáticos. Penetran en la medula espinal por medio de
cuatro o cinco raíces dorsales torácicas
superiores.
24. DEFINICION
DE MICROCIRCULACION.- IDENTIFIQUE LOS COMPONENTES HISTOLOGICOS DE
LA MICROCIRCULACION.
La
microcirculación es una parte del sistema circulatorio
encargada del intercambio de gases, fluidos, nutrientes y
productos metabólicos de desecho.
A este nivel
los fenómenos de regeneración y permeabilidad son
máximos.
La microcirculación es el flujo de sangre que
discurre por vasos sanguíneos de 100 micras de diámetro
o menos (p.e. arteriolas, capilares y vénulas). Las
principales funciones de la
microcirculación son el transporte de sangre hacia y
desde los tejidos, y servir como líquido refrigerante del
cuerpo en procesos de termorregulación. También
contribuye al color y la turgencia de los tejidos.
ARTERIOLAS: Son las ramas terminales del árbol que
irrigan el lecho capilar.
Tamaño estimado: menor que 0,1 mm.
Existen una
transición gradual en la estructura y la función entre
estos tres tipos de vasos arteriales, más que una
demarcación abrupta. En general, la cantidad de tejido
elástico disminuye según los vasos se hacen más
pequeños y el componente de tejido muscular liso asume una
preponderancia relativamente mayor.
Son
estructuras vasculares más pequeñas con respecto a las
arterias musculares. Vemos menos células endoteliales,
proporcionales a la superficie endotelial (se reduce el
lúmen). Los núcleos de las células endoteliales
hacen cierta prominencia hacia la luz del pequeño vaso.
Presentan una capa media gruesa, muscular con 4 a 5 capas de
músculo liso. Por fuera, hacia la adventicia hallamos un
tejido de sostén conformado por abundantes fibras
colágenas que delimitan claramente la adventicia de la
media.
La
función principal de estos vasos arteriales es la de
control del flujo
sanguíneo al territorio capilar (vasos de resistencia
periférica).
En las
arteriolas pequeñas hallamos de 1 a 2 capas de fibras
musculares lisas, concéntricas con una lámina
elástica interna y una adventicia bastante desarrollada.
Características Histológicas:
A nivel de la
Túnica íntima, con una capa subendotelial más
fina, sin lámina elástica limitante interna (la cual
podemos encontrar en arteriolas de mayor calibre).
La
Túnica Media, conformada por 1 a 2 capas de células
musculares lisas, sin encontrarse limitante externa de naturaleza elástica.
La
Túnica Adventicia se halla poco desarrollada.
En vasos
sanguíneos que poseen músculo liso (arteriolas y
arterias musculares, las células endoteliales poseen
frecuentes prolongaciones que atraviesan la lámina basal y
la elástica interna (cuando existe), entrando en contacto
directo con las células musculares lisas de la media.
De las
arteriolas en adelante, es decir disminuyendo el diámetro de
la luz nos adentramos en el terreno de la
microcirculación.
Las
arteriolas más pequeñas presentan una sola célula
endotelial, posee escaso músculo liso y su lúmen puede
llegar a 5 micrones o menos. Cuando el músculo liso presente
en la pared se contrae el lúmen puede obliterarse casi
completamente. Esta arteriola se llama metaarteriola.
Cuando
la metaarteriola se ramifica, el músculo liso ya no se
observa continuo sino que se hace "intermitente", llegando a
convertirse en una arteriola precapilar.
CAPILARES:
Son vasos más pequeños, de paredes muy delgadas. Su
lúmen no suele ser mayor que 7 micrones. Los glóbulos
rojos pasan como tapones o émbolos por el lúmen de
estos pequeños vasos.
q Capilares Fenestrados: Las células
endoteliales poseen numerosos poros.
En estos
capilares encontramos verdaderos "poros", aberturas o
fenestraciones. Las fenestraciones se hallan limitadas por la
presencia de membrana basal. Estos "poros" tienen un calibre
determinado (90 nm de diámetro). La filtración se
presenta principalmente a nivel de las fenestraciones. La
mayoría de los "poros" poseen diafragmas, los diafragmas
serían selectivos al regular el paso de distintas
sustancias. Cada diafragma tiene un diámetro aproximado de
80 nm. Este tipo de capilares fenestrados los encontramos en el
estómago, intestino delgado, etc. La presencia de poros
determinaría que la cantidad de vesículas
pinocíticas generadas por la célula endotelial
sería menor con respecto a los capilares continuos.
¿Dónde los encontramos?: Glándulas
endocrinas y sitios de absorción de líquidos y
metabolitos tales como vesícula biliar, tubo digestivo.
q Capilares discontinuos o sinusoidales
(sinusoides): A diferencia de los capilares anteriores, este
tipo de capilar son mucho más anchos, con diámetros
variables (30 – 70 micrones). El trayecto de estos capilares es
habitualmente "tortuoso", siguiendo o adaptándose a la forma
de los espacios que van dejando las células con las cuales
se relaciona estrechamente. La pared de éstos capilares
puede ser continua o bien fenestrada. La lámina basal
presente es incompleta.
Encontramos
capilares sinusoidales en el hígado, médula ósea y
Bazo.
Endotelio discontinuo: Las células endoteliales no
forman una interfase continua entre la luz y tejidos
circundantes; esta disposición se encuentra sólo en los
sinusoides del hígado, bazo y médula ósea.
En los
capilares continuos y fenestrados, la filtración comprende
principalmente líquido tisular. En los capilares
discontinuos no sólo hay intercambio de metabolitos; sino
también de células (aquí los capilares en lugar de
medir 7 micrones de diámetro, los hallamos de 30 micrones).
Las células endoteliales son alargadas, hallándose
grandes espacios entre ellas. Casi no presentan membrana basal.
Conservan su disposición gracias a la presencia de fibras
reticulares. Las células endoteliales se observan unidas
sólo en ciertos puntos. En muchos puntos existen verdaderas
"ventanas", que en ocasiones pueden ser ocluidas por células
con capacidad fagocítica.
¿Dónde los encontramos?: Hígado, Bazo,
Médula ósea.
q Presencia de "shunt" o cortocircuitos
arterio-venosos. Proporcionan conexiones directas entre el
sistema arterial y venoso.
q La abundancia de la red capilar. Depende de los
requerimientos funcionales del tejido. Por ejemplo, el tejido
colágeno denso de los tendones tiene una red de capilares
dispersa; en contraste, el músculo cardiaco tiene una
extensa red de capilares que se extiende por el intersticio entre
las fibras musculares.
No olvidemos que el control de la red capilar se origina en
las propiedades contráctiles del músculo liso, que
regula el flujo sanguíneo terminal a través de 4
sistemas:
El nervioso autónomo; el sistema de control por
sustancias químicas (histamina, noradrenalina);
Sistema endocrino (hormonas) y la hipoxia
celular (contracción de oxígeno).
SISTEMAS VASCULARES ESPECIALES:
SISTEMAS DE VASOS PORTA.
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |