- Introducción
- Definición de tejido
muscular - Musculo estriado o
esquelético - Músculo
cardíaco - Músculo liso
- Contracción muscular

Introducción
Ya en 1863 Sechenov decía que toda
manifestación externa del ser vivo es la actividad
muscular. Exceptuando la emisión de luz por los seres
vivos (bioluminiscencia), la contracción muscular es
realmente el medio de manifestación de actos tanto
internos como externos. La contracción muscular deriva de
la actividad cerebral, pero sin ella todas las conquistas
intelectuales permanecerían confinadas al sistema
nervioso.
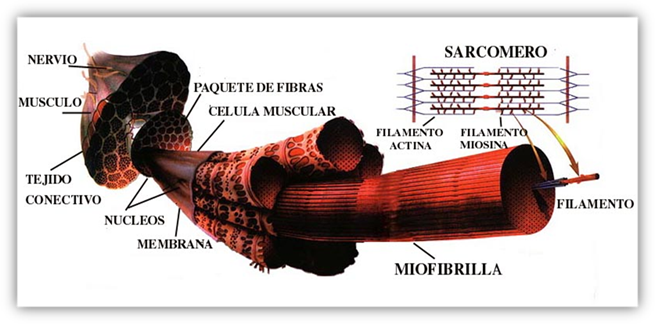
Todo músculo está formado por un haz de
fibras. Existen dos tipos principales de fibras musculares, las
lisas y las estriadas. Esa nomenclatura viene del aspecto
microscópico.Las fibras lisas se contraen más
lentamente, pero la contractura puede durar mucho tiempo. Son
encontradas en las vísceras, especialmente en el tubo
digestivo, vejiga y arterias. Las fibras estriadas se contraen
más rápidamente y en casos normales su
contracción dura poco. Forman las masas de los
músculos esqueléticos, y como un tipo especial del
miocardio. La musculatura estriada representa el 40% de las masa
corporal humana. Existen ciertos músculos lamelibranquios
que poseen un músculo con dos tipos de fibras: las
estriadas cierran rápidamente la concha pero con poca
fuerza. Las fibras lisas mantienen una contracción fuerte
por un largo periodo. Ese comportamiento es importante en la
defensa del molusco.
La fibra muscular genera tensión a través
de la acción de la actina y la miosina. Mientras
está en tensión, el músculo puede alargarse,
acortarse, o permanecer igual. Aunque el término
contracción implica acortamiento, cuando se hace
referencia al sistema muscular, significa fibras musculares
generando de tensión con la ayuda de las neuronas motoras.
Estas contracciones producen la fuerza motora de casi todos los
músculos superiores, como por ejemplo, desplazar el
contenido de la cavidad a la que recubren (músculo liso) o
mover el organismo u otros objetos a través del medio
(músculo estriado). Las contracciones involuntarias son
controladas por el sistema nervioso central, mientras que
el cerebro controla las contracciones voluntarias, y
la médula espinal controla
los reflejos involuntarios.
Definición de
tejido muscular
Es un Tejido CONTRÁCTIL formado por
células llamadas fibras musculares que ejercen
tracción mediante tendones sobre un sistema de palancas
articuladas (huesos y articulaciones). El tejido muscular se
especializa en producir movimientos, tanto del cuerpo en conjunto
como de sus partes entre sí. Las células musculares
muestran gran desarrollo de la función de contractilidad,
y en menor grado de la conductividad. Esta especialización
incluye el alargamiento de las células en el eje de
contracción, y a causa de esto las células se
llaman a
menudo fibras musculares.
En el tejido muscular, las células o fibras
musculares suelen agruparse en haces, pero el tejido muscular no
sólo consta de fibras musculares. Las fibras musculares,
por realizar trabajo mecánico, necesitan una red abundante
de capilares sanguíneos que les proporcionen materiales
alimenticios y oxígeno, y eliminen los productos
tóxicos de desecho. Los vasos sanguíneos son
conducidos por el tejido fibroconectivo, que también sirve
para mantener unidas las fibras musculares y para proporcionarles
el sostén necesario para que puedan ejercer su
tracción con utilidad. Los nervios también cursan
en el tejido conectivo. Hay tres tipos de músculo,
que se clasifican sobre bases estructurales y funcionales. Hay
bandas transversales regulares a lo largo de las fibras del
músculo estriado, mismas que faltan en el
músculo liso. En el aspecto funcional, el
músculo puede estar bajo control de la mente
(músculo voluntario) o no estarlo
(músculo involuntario). Los tres tipos de
músculo son:

Para el estudio histológico del
tejido muscular (con microscopio óptico y
electrónico) se usa principalmente material en cortes,
pero se puede obtener información adicional útil
del tejido macerado. En este método se
sumergen fragmentos pequeños de músculo en una
solución a 10% de ácido clorhídrico en
solución salina fisiológica por 24 a 48 horas.
Después de lavar con agua, las fibras musculares se pueden
disgregar en un cubreobjetos usando dos agujas montadas, luego se
cubren con un cubreobjetos y se examinan al
microscopio.
Musculo estriado o
esquelético
El músculo estriado o esquelético es el
que forma la carne del humano. En estado fresco tiene color rosa,
debido en parte a un pigmento que hay en las fibras musculares, y
en parte a la abundante vascularización del tejido; sin
embargo, hay alguna variación en el color, y se han
identificado músculos "rojos" y "blancos". La
célula o fibra muscular individual es larga,
cilíndrica y multinucleada, con los extremos terminados en
punta o algo redondeados en la unión entre músculo
y tendón. Cada fibra es independiente y puede ser de gran
longitud, pero en muchos músculos las fibras individuales
son más cortas que la longitud total del músculo,
con un extremo unido a un tendón y el otro a un tabique de
tejido conectivo en el interior del
músculo.
En general la fuerza de un músculo
depende no sólo de la longitud de sus fibras musculares,
sino también del número total de fibras que hay en
él. Con el ejercicio, los músculos aumentan de
volumen, y esto se debe a un aumento de volumen de cada fibra
muscular individual (hipertrofia) y no al
aumento del número de
fibras (hiperplasia). La contracción
del músculo estriado es más rápida que la
del músculo liso. Cada uno de los llamados
músculos en anatomía macroscópica
está envuelto por una capa de tejido conectivo
relativamente grueso llamado epimisio, que a
simple vista aparece como una vaina blanca. Dentro de él
están las fibras musculares dispuestas en haces o
fascículos. Cada fascículo está rodeado por
una vaina de tejido conectivo más delgado,
el perimisio. Dentro de un fascículo,
las fibras musculares individuales están cubiertas por
tejido conectivo fino,
el endomisio.
En las preparaciones por
disgregación de músculo fresco o macerado, las
fibras individuales tienen color amarillo pálido. Las
fibras son estriadas en sentido longitudinal así como
transversal y varían de 1 a 40mm de longitud y de l0 a
100um de diámetro. En cada fibra los núcleos son
numerosos (alrededor de 35 por mm de longitud), ovoides y
situados en la periferia, adyacentes al sarcolema. Este es una
membrana delgada sin estructura que limita a la fibra, y que se
ve con claridad en las regiones en que la fibra se ha aplastado,
con la consiguiente retracción del sarcoplasma dentro del
sarcolema.
En las preparaciones sistemáticas,
el sarcoplasma está ocupado principalmente por haces
filamentosos largos, cilíndricos, paralelos, de una a tres
micras de diámetro. Estas son
las miofibrillas, que muestran
estriación transversal. En los cortes transversales
aparecen como puntos con citoplasma claro no fibrilar entre ellas
y rodeándolas. También se encuentra sarcoplasma
claro en zonas adyacentes a los núcleos, y en estos
lugares contiene numerosos sarcosomas (mitocondrias),
pequeños aparatos de Golgi, algunas gotitas de
lípido y glucógeno. Hay más glucógeno
y sarcosomas entre las miofibrillas. Los sarcosomas
interfibrilares suelen mostrar disposición regular en
relación con la estriación transversal de las
miofibrillas, encontrándose entre pares de bandas 1
adyacentes, a ambos lados de la banda Z, con otros sarcosomas
dispuestos en hileras longitudinales y acúmulos bajo el
sarcolema.


Características del
Músculo Estriado

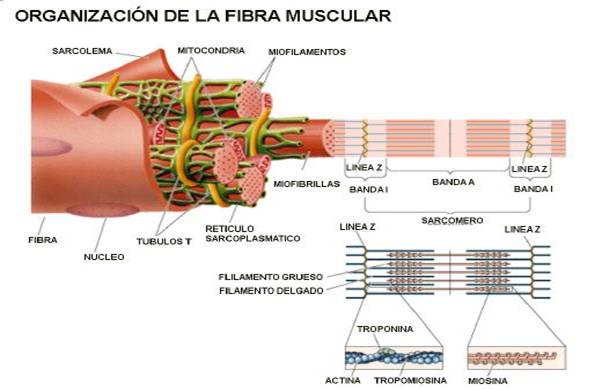
3.1.1Miofibrillas y Estriación
Al microscopio óptico, la fibra muscular en corte
longitudinal muestra bandas alternas oscuras (A) y claras (1).
Con luz polarizada, las bandas A son birrefringentes
o anisotrópicas, y se ven brillantes;
las bandas I aparecen oscuras y sonisotrópicas, de
aquí el nombre de las bandas. Cada banda A muestra una
zona central menos birrefringente llamada banda H, y cada banda 1
está dividida por una línea o banda Z bien
definida. Aunque las bandas parecen cruzar por completo la fibra
muscular, en una buena preparación se puede ver que
están limitadas a las miofibrillas y que no se ven en el
sarcoplasma interfibrilar. El segmento entre dos
líneas Z adyacentes se
denomina sarcómera, de unas 2 a 3um de
longitud, de las cuales 1.5 m corresponden a la banda A y 0.8 a
la banda I. La sarcómera no sólo es una unidad
estructural, sino también la unidad contráctil
básica. En el músculo relajado en corte
longitudinal las bandas están bien marcadas. Durante la
contracción, las miofibrillas son más gruesas y las
sarcómeras más cortas, ya que la distancia entre
las líneas Z se acorta en forma progresiva. A medida que
las bandas I se hacen más cortas, los extremos de las
bandas A se aproximan a las líneas Z, hasta que al final,
en contracción total, las bandas A e I son
indistinguibles; pero la longitud de la batida A en la
contracción permanece constante. Estas
bandas no son visibles en el corte transversal de una fibra
muscular estriada.
3.1.2 Estructura Fina
Las miofibrillas, que aparecen como
filamentos largos y paralelos, de una a 3um de diámetro al
microscopio óptico, están formadas por unidades
más pequeñas
llamadas miofilamentos. Hay dos tipos
principales de ellos según sus calibres (gruesos y
delgados) y su composición química. Los dos tipos
están dispuestos de manera regular, y las bandas
transversales que se ven al microscopio óptico sólo
son un reflejo de su distribución en la sarcómera.
Los filamentos gruesos contienen principalmente miosina. Miden
alrededor de 12 a 15 nm de diámetro y 1.5um de longitud, y
se encuentran en el centro de la sarcómera ocupando la
banda A. Los filamentos delgados contienen actina, sólo
miden 5nm de diámetro y 1 1um de longitud y se extienden
de cada lado de la banda Z y pasan a través de la banda 1
adyacente y parte de la banda A, donde se interdigitan con los
filamentos gruesos, por lo que en corte transversal de la banda
A, cada filamento grueso está rodeado por seis de los
delgados en disposición hexagonal. La banda H sólo
es la zona central de la banda A y está libre de
filamentos delgados, y su anchura está determinada por el
grado de contracción. Así, el diseño de las
bandas se explica por la presencia o ausencia de
superposición entre los dos tipos de filamentos. Por
tanto, en corte transversal la banda 1 sólo contiene
filamentos delgados y las extremidades de la banda A contienen
gruesos y delgados. En la banda H sólo hay filamentos
gruesos. En la línea M al centro de ella, los filamentos
gruesos están interconectados por filamentos finos
dispuestos en forma radial, de manera que cada uno está
conectado en el plano transversal con los seis filamentos
adyacentes. Es probable que estos filamentos transversales de la
línea M ayuden a mantener la disposición y el
espaciamiento regulares de los filamentos gruesos en la
sarcómera. En las regiones de superposición
(esto es, en las extremidades de la banda A), cada filamento
grueso de miosina está rodeado por seis filamentos
delgados de actina. El filamento grueso muestra estructuras a
manera de puentes que se extienden en forma radial hacia los
filamentos delgados. Estos puentes faltan en el centro de la
banda H.
Un filamento grueso está formado por un haz
de moléculas de miosina, cada una en forma de palo de golf
con un tallo y una cabeza. La molécula de miosina tiene
dos subunidades, la neromiosina ligera, que forma la mayor parte
del tallo, y la meromiosina pesada, que forma el resto del tallo
y la cabeza. Son las cabezas de las moléculas las que
sobresalen del haz para extenderse como puentes transversales
entre los filamentos gruesos y los delgados.
Las cabezas se disponen en espiral alrededor del
filamento grueso, de manera que se forman seis hileras
longitudinales que conectan con los seis filamentos delgados
relacionados y el filamento grueso está polarizado, ya que
as cabezas están dirigidas alejándose de su parte
media (en la línea M). En la parte central del haz
sólo se observan os tallos de las moléculas. La
cabeza de la molécula de miosina es flexible en el tallo,
y las cabezas o puentes son los que poseen actividad de
adenosintrifosfatasa (ATPasa) necesaria para la
interacción de la actina y la miosina durante la
contracción.
Un filamento delgado está formado
principalmente por F-actina, una proteína filamentosa
compuesta de dos hilos de subunidades globulares de G-actina,
hilos que están enrollados de manera helicoidal. La
F-aetina tiene polaridad lineal; todos los filamentos que se
insertan en un lado de la línea Z son
de la misma polaridad, y los que se insertan ene1 otro lado de la
línea Z (y en el otro extremo de la misma
sarcómera) son de polaridad opuesta. En los filamentos
delgados también hay tropomiosina y troponina. Las
moléculas de tropomiosina son largas y delgadas y se
encuentran de un extremo a otro en los surcos de la doble
hélice de F-actina; las moléculas de troponina
globular están unidas a la tropomiosina a intervalos
regulares. En la línea Z, los filamentos delgados de
sarcómera adyacentes no están en registro; esto da
un aspecto característico de zigzag a la línea
Z.
Parece que cada filamento delgado de una
sarcómera está unido por filamentos muy finos a
cuatro filamentos delgados de la sarcómera adyacente;
estos filamentos finos forman un diseño tetragonal en el
corte transversal. Los componentes químicos de la
línea Z incluyen actina, tropomiosina y alfa-actinina,
pero su composición estructural aún no es muy
clara. Dado que los filamentos delgados miden 1.0um de
longitud y los filamentos gruesos miden 1.5um, es obvio que
cuando la longitud de la sarcómera sea menor de 2um, los
filamentos delgados originados en líneas Z adyacentes se
tocarán en el centro de la banda A, y que una mayor
contracción hará que los extremos de los filamentos
gruesos se pongan en contacto con las líneas Z cuando la
longitud de la sarcómera llegue a 1.5um. En las
células musculares hay otros filamentos aparte de los
miofilamentos delgados y gruesos. Los filamentos
intermedios (10 nm) forman una red extensa. En un plano
transversal, hay fascículos de filamentos
transversales cortosque conectan miofibrillas adyacentes,
pasando entre sus líneas Z y M. En un plano longitudinal,
hay filamentos continuos, largos y paralelos, que
conectan la periferia de líneas Z sucesivas para formar
una vaina a la sarcómera. Se cree que estos filamentos
desempeñan un papel importante como citoesqueleto en la
estructura y la mecánica de la fibra muscular.
3.1.3 Sistemas de Membranas
El sarcolema, demasiado delgado para ser observado
con claridad con el microscopio óptico, consta de la
membrana plasmática de la célula muscular cubierta
por una lámina basal extracelular fina con algunas
microfibrillas de colágena relacionadas con ellas. Entre
las fibras musculares también se encuentran bandas
delgadas de tejido elástico.
El retículo sarcoplásmico representa
un tipo especial de retículo endoplásmico agranular
(liso). Es extenso y comprende un sistema continuo de
túbulos y cisternas limitados por membrana que forman una
vaina o collar alrededor de cada miofibrilla, con comunicaciones
entre los collares de sarcómeras adyacentes de
miofibrillas vecinas. En el músculo de los
mamíferos, la vaina está formada principalmente por
elementos tubulares en disposición longitudinal alrededor
de la banda A, con comunicaciones transversales que son notables
en la región de la banda H. En las uniones A-I hacia los
extremos de las sarcómera, los túbulos están
unidos a las cisternas, llamadas cisternas terminales, que tienen
forma de anillos alrededor de las miofibrillas en las uniones
A-I. Una red semejante de túbulos rodea las bandas 1, con
otra cisterna terminal en cada unión A-I, pero
están separados por un tú- bulo transversal
más delgado llamado túbulo T.Esta
disposición de dos cisternas terminales del
retículo sarcoplásmico y un túbulo T central
se llama tríada, y hay dos
tríadas por cada sarcómera en el músculo de
los mamíferos. (En el músculo de los anfibios, una
triada rodea a cada miofibrilla en la línea Z, o sea que
las tríadas se encuentran entre las
sarcómera.)
Los túbulos transversales (T) son
invaginaciones del sarcolema superficial, y su luz se
continúa con el espacio extracelular. Estos pasan del
sarcolema al interior de la fibra muscular a intervalos
regulares, se ramifican y, como ya se señaló, se
encuentran entre las cisternas terminales del retículo
sarcoplásmico para formar las tríadas en las
uniones A-I. En las tríadas, las membranas de los
túbulos T y del retículo sarcoplásmico
forman uniones o acoplamientos. Otras
características del sarcoplasma que se ven por microscopia
electrónica incluyen la presencia de numerosos sarcosomas
o mitocondrias. Estas por lo general son grandes, con crestas
agrupadas muy juntas, y se encuentran subyacentes al sarcolema,
concentradas en los polos nucleares y en hileras paralelas
dispuestas de manera regular entre las miofibrillas. El
sarcoplasma paranuclear suele contener un pequeño aparato
de Golgi, algunos ribosomas y unos cuantos elementos del
retículo endoplásmico granular. También se
pueden observar algunos lisosomas y numerosas partículas
de glucógenos dispersas por toda la célula, pero a
menudo concentrado entre las miofibrillas a nivel de las bandas
I. Normalmente hay vacuolas de lípido que pueden aumentar
con la edad.
3.1.4 Contracción
En la contracción, las fibras musculares se hacen
más cortas y anchas. Estudios con microscopio de contraste
de fase y de interferencia demuestran que durante la
contracción la longitud de la banda A permanece constante,
en tanto que la de las bandas 1 y H disminuyen, aunque sin
alterarse la de los filamentos, ni gruesos ni delgados. La
contracción se realiza por el mecanismo del "filamento
deslizante", que entraña un cambio en la posición
de las dos series de miofilamentos; los filamentos delgados se
deslizan más allá de los gruesos y son
atraídos hacia adentro de la sarcómera, hacia la
línea M. Esto hace que las líneas Z se acerquen
entre sí, y así se acorta la sarcómera, con
disminución de la anchura de las bandas 1 y H y,
finalmente, su eliminación en la contracción
completa, cuando los extremos de los filamentos gruesos se
aproximan a las líneas Z. En el mecanismo de la
contracción, las cabezas de las moléculas de
miosina se desplazan más que las subunidades globulares de
actina, se unen con ellas, y por medio de un ciclo de trabado,
destrabado y retrabado se genera la fuerza contráctil,
deslizándose los filamentos unos sobre otros y atrayendo
los filamentos delgados hacia adentro a manera de una rueda
dentada animada. Este fenómeno necesita la energía
que deriva del desdoblamiento del adenosintrifosfato (ATP) en
adenosindifosfato (ADP) y fosfato, en que las cabezas de miosina
actúan como una ATPasa. La reserva de ATP es limitada, y
se re- abastece de continuo por los sarcosomas que transportan as
enzimas del ciclo del ácido cítrico, y las dos
series de filamentos se deslizan unos sobre otros de regreso a su
posición original de reposo, en que están
parcialmente superpuestos.
El sistema T y el retículo sarcoplásmico
también participan en la contracción. Al estimular
una fibra muscular, todas las miofibrillas se contraen de manera
simultánea e instantánea. La contracción
empieza en la unión A-I (en el lugar de una tríada)
y se sabe que los iones calcio son esenciales para esto. En el
músculo relajado, la mayor parte del calcio está
concentrado en el retículo sarcoplásmico. Al
estimular una fibra muscular, una onda de despolarización
de la membrana pasa rápidamente sobre la membrana
plasmática y se extiende al interior por el sistema T.
Esto produce un cambio en la permeabilidad de las cisternas
terminales del retículo sarcoplásmico con
liberación de calcio hacia el sarcoplasma que rodea
las miofibrillas. Luego el calcio se une a la troponina, que a su
vez ocasiona que la molécula de tropomiosina se profundice
más en el surco de la doble hélice de F-actina,
para así descubrir los sitios del filamento de actina que
estaban "bloqueados" previamente. Estos sitios quedan ahora
disponibles para el contacto con las cabezas de miosina.
Así, el calcio "abre" los sitios de unión activa
con la miosina en el filamento delgado y permite que las cabezas
de miosina giren y se unan a ellos para iniciar el mecanismo de
la contracción.
Después del estímulo se invierte la
secuencia: el calcio regresa al retículo
sarcoplásmico y las moléculas de tropomiosina
regresan a una posición más periférica a lo
largo de los filamentos delgados, bloqueando los sitios activos
de la actina. En el músculo relajado, las moléculas
de troponina y tropomiosina (las "proteínas reguladoras")
forman un mecanismo de cierre que evita la interacción
miosina-actina. Durante la contracción, el calcio es la
"llave" que abre este mecanismo. Hay que hacer notar que una
célula del músculo esquelético sólo
puede contraerse a su máxima capacidad, la "ley del todo o
nada", y que la fuerza de la contracción varía
según el número de unidades musculares que se
contraen.
3.1.5 Tipos de fibras Musculares
Desde el punto de vista macroscópico, los
músculos difieren en su color, pues algunos son más
rojos que otros. Estos músculos están formados por
tipos diferentes de fibras musculares.
Las fibras que predominan en los llamados
músculos "rojos" son las fibras
rojas, que tienen un diámetro relativamente
pequeño, con numerosos sarcosomas grandes que muestran
sus crestas agrupadas muy juntas, y que se disponen en
acúmulos bajo el sarcoplasma y en hileras
longitudinales entre las miofibrillas.Las fibras blancas son más
grandes y forman la masa de los músculos "blancos",
sus sarcomas son más pequeños y se encuentran
principalmente por pares alrededor de las líneas Z,
que sólo tienen la mitad de la anchura de las que se
observan en las fibras rojas.Las fibras intermedias son semejantes
alas rojas; se encuentran en los músculos rojos, pero
tienen sarcosomas más pequeños y una
línea Z más delgada.
Las uniones mioneurales son más complejas en las
fibras blancas, y la distribución de los tipos de fibras
en un músculo parece estar bajo la influencia del sistema
nervioso. Las fibras rojas se contraen con más lentitud
que las blancas y se fatigan con menor rapidez, aunque de hecho
hay dos tipos de fibras rojas, uno de los cuales se contrae muy
rápidamente. Aunque en el aspecto morfológico las
fibras intermedias son semejantes a las fibras rojas, en la
velocidad de su contracción se asemejan más a las
fibras blancas. Algunos músculos contienen los cuatro
tipos de fibras musculares, cada uno con diferente tiempo de
contracción.
3.1.6 Organización
El tejido conectivo relacionado con las fibras
musculares tiene tres grados de grosor:
Todo el músculo está envuelto por una
capa consistente de tejido conectivo,
el epimisio.Las divisiones más delgadas de tejido
conectivo que rodean a los haces o fascículos de
fibras musculares forman el perimisio.Hay tejido conectivo aún más delgado
que se extiende a partir del perimisio para penetrar entre
las fibras y rodearlas
como endomisio. Este consta de una red de
fibras reticulares y algunas células de tejido
conectivo, y una cantidad variable de fibras elásticas
(particularmente notables en los músculos
pequeños del ojo y de la cara). En el endomisio se
encuentran capilares sanguíneos y nervios
pequeños.
3.1.7 Riego sanguíneo y
linfático
El músculo estriado dispone de un riego
sanguíneo muy rico. Las arterias provienen del epimisio y
siguiendo el perimisio penetran en la sustancia del
músculo. Las arterias se ramifican en arteriolas y
terminan en capilares, que siguen por las finas trabéculas
del endomisio. La mayor parte de capilares se hayan dispuestos
paralelamente a las fibras musculares, pero es frecuente que
desprendan ramas en ángulo recto.
Casi todos los linfáticos del músculo
estriado se hayan en su estructura conectivas más gruesas.
En este punto difieren del músculo cardiaco, que tiene
linfáticos y capilares entre las fibras.Los vasos
linfáticos que drenan el líquido tisular son muy
numerosos, pero no se encuentran en relación íntima
con las fibras. Se encuentran en el epimisio y el perimisio, pero
no en el endomisio.
3.1.8 Innervación
Los nervios penetran en los músculos siguiendo el
tejido conectivo de estos últimos. El número de
fibras musculares inervadas por una sola fibra nerviosa es muy
variable. En uno de los músculos extrínsecos del
ojo, donde se requiere una extraordinaria presión en los
movimientos, hay una fibra nerviosa para cada fibra muscular. El
extremo opuesto está representado por los músculos
que no han de efectuar movimientos delicados, en los cuales una
sola fibra nerviosa puede ramificarse e inervar más de un
centenar de fibras musculares. Una fibra nerviosa y las fibras
musculares por ella inervadas reciben en conjunto el nombre de
unidad motora. Si una sola fibra inerva varias fibras musculares
de un músculo, estas no constituyen un grupo localizado
como pudiera pensarse; por lo contrario, las fibras musculares
inervadas por una sola fibra nerviosa puede estar distribuidas
muy difusamente por todo el músculo. Sin embargo son todas
rojas o todas blancas. Estos descubrimientos tienen importancia
porque una fibra muscular aislada, bajo la influencia de un
estímulo nervioso, siempre se contrae a su capacidad
máxima; se dice que esto constituye la llamada "ley del
todo y nada". En consecuencia, la capacidad de un músculo
en conjunto para contraerse con diversas intensidades no dependen
de que cada una de las fibras musculares se contraigan con
intensidad diversa, cosa imposible, sino de que pueda estimularse
un numero diferente de fibras que se contraigan. Por lo tanto si
se requiere una contracción débil, solo con
estimuladas y obligadas a contraerse una pequeña
proporción de las fibras del músculo. En tales
condiciones, interesa que las que entren en acción
representen el músculo en conjunto para que la
contracción resulte general y no local. Como muchas fibras
pueden estar inervadas por una sola fibra nerviosa, y como en las
contracciones poco intensas solo se estimula una
proporción muy pequeñas de fibra, interesa que cada
fibra inerve un grupo de fibras musculares ampliamente
distribuidas a todo lo largo del músculo.
3.1.9 Placa Motora o Terminal
Cada fibra muscular tiene una placa motora Terminal
o unión neuromuscular (mioneural), o
sea el lugar en que una terminación nerviosa
(terminación axónica) se relaciona
íntimamente con una fibra muscular. Cuando el axón
se aproxima a la fibra muscular, pierde su vaina de mielina, pero
permanece cubierto por una vaina delgada de citoplasma de la
célula de Schwann. La terminación nerviosa se
ramifica para formar una masa semejante a una placa o un conjunto
de abultamientos terminales como un racimo de uvas, que se
sitúan en una hendidura de la superficie muscular. Estas
características se demuestran por microscopia
óptica con técnicas de impregnación
argéntica. Por microscopia electrónica se observa
que los abultamientos terminales contienen numerosas
vesículas sinápticas pequeñas que contienen
acetilcolina y algunas mitocondrias, pero no filamentos ni
microtúbulos (éstos son notables más
proximalmente en el axón). En el lugar de la unión,
el sarcolema forma una depresión poco profunda (la
hendidura sináptica primaria) en que se encuentra la
terminación axónica, y después se invagina
para formar hendiduras secundarías profundas o pliegues de
unión. Entre la terminación nerviosa y el sarcolema
hay una lámina basal sencilla formada por la fusión
de la del sarcolema, el axón y su célula de
Schwann. Esta lámina basal se extiende hasta los pliegues
de unión. En la placa motora Terminal, la fibra muscular
muestra núcleos y numerosos sarcosomas en la región
llamada sarcoplasina de unión.
Funcionalmente, y al parecer en relación con la
lámina basal, la hendidura
sináptica, que sólo tiene 20 a 50 nm de
ancho, contiene acetilcolinesterasa, la enzima que inactiva al
neurotransmisor acetilcolina. Esta última es liberada por
las vesículas sinápticas por exocitosis, pasa a
través de la hendidura sináptica hasta sitios
receptores del sarcolema localizados principalmente en las partes
altas de los pliegues de unión y produce la
despolarización del sarcolema. Después de la
liberación de acetilcolina, la membrana de la
vesícula se reincorpora al axón por endocitosis.
Después de ser liberada, la acetilcolina es degradada con
rapidez por la acetilcolinesterasa para permitir estímulos
repetidos posteriores. y de las cuales hay dos tipos en los
músculos:
1. Husos neuromusculares. Son
fusiformes, están situados en forma longitudinal en el
músculo y se forman por varias fibras musculares
pequeñas y delgadas, las fibras
intrafusales, envueltas por una cápsula de
tejido conectivo con la que se relaciona una terminación
nerviosa. Las fibras intrafusales son de dos tipos: las primeras,
las fibras de bolsa nuclear,son más grandes
y en menor número, con muchos núcleos concentrados
en el centro de la célula; las segundas, las fibras de
cadena nuclear, son más pequeñas y numerosas y
contienen los núcleos en una hilera. Las fibras nerviosas
aferentes terminan en las fibras de bolsa nuclear formando una
espiral (terminaciones anuloespirales) y en las fibras de cadena
nuclear como racimos (terminaciones en ramillete); estas fibras
también tienen inervación motora. Las terminaciones
sensoriales son propioceptivas y responden a la
distensión, por lo que participan en los reflejos de
estiramiento. 2. Las terminaciones
neurotendinosas (órganos neurotendinosos) se
encuentran en los tendones cerca de la unión
musculotendinosa y muestran fibras nerviosas sensoriales que
terminan entre los haces de fibras tendinosas (colágenas).
Estas fibras son estimuladas por la tensión o el
estiramiento del tendón durante la contracción
muscular.
3.1.10 Regeneración
Después de una lesión, las fibras
musculares degeneradas tienen capacidad limitada de
regeneración, pero el daño macroscópico se
repara por medio de tejido conectivo fibroso, por lo que se forma
una cicatriz. De manera semejante, si se interrumpe la
innervación o el riego sanguíneo, las fibras
musculares degeneran y son sustituidas por tejido fibroso. Sin
embargo en el músculo del adulto hay células
satélites. Estas pequeñas células
uninucleadas se encuentran entre el sarcolema y el endomisio y es
probable que representen un reservorio de mioblastos
embrionarios. Durante la vida pueden definirse y participar en la
separación y regeneración que pueden presentarse en
el músculo maduro.
Músculo
cardíaco
Aunque es estriado es involuntario, se contrae de manera
rítmica y automática. Se encuentra sólo en
el miocardio (capa muscular del corazón) y en las paredes
de los grandes vasos unidos a éste órgano. Una
fibra del músculo cardíaco es una unidad lineal
formada por varias células musculares cardíacas
unidas por todos sus extremos en zonas especializadas de
unión llamadas "discos intercalados". Cada célula
mide alrededor de 100um de longitud y 15um de diámetro; a
menudo divididas parcialmente en dos ó más ramas en
sus extremos, ramas que se unen a células adyacentes, o
partes de ellas en los discos intercalados.
El aspecto global es el de fibras principalmente
paralelas con numerosos haces cruzados, que da la falsa
impresión de ser una red sinticial. Entre las fibras, hay
tejido conectivo fino, en el endomisio, que contiene los vasos
sanguíneos y linfáticos pequeños.La fibra
del músculo cardíaco está envuelta por un
sarcolema delgado y tiene sarcoplasma abundante con numerosas
mitocondrias dispuestas en hileras entre ellas, y en consecuencia
hay una obvia estriación longitudinal. También se
observa un modelo de seriación transversal de las
miofibrillas. Los núcleos son alargados y están
situados en el centro de la fibra entre miofibrillas divergentes.
Alrededor de los núcleos hay áreas fusiformes de
sarcoplasma que contienen muchas mitocondrias, un pequeño
aparto de Golgi en un polo del núcleo, algunas gotitas de
lípidos y, al aumentar la edad, algunos depósitos
del pigmento lipofucsina (lisosomas secundarios); el cual puede
extenderse hasta el grado de darle un tinte parduzco al miocardio
recién obtenido, lo que se conoce como "atrofia parda del
corazón".
4.1 Histogénesis
La musculatura cardíaca evoluciona a partir de
mioblastos que difunden desde la porción del mesodermo
esplácnico que rodea los tubos cardiacos limitados por
endotelio. Las células continúan su división
por mitosis durante y después de finalizada la
diferenciación hasta poco antes del nacimiento, lo cual
indica condiciones bien distintas de las que rodean el desarrollo
de la musculatura esquelética, en la cual las
células ya diferenciadas ya no se dividen, ni siquiera
durante la vida fetal.
4.2 Estructura Fina
Los miofilamentos que contienen actina y miosina
están limitados a las células musculares
individuales y no cruzan las uniones celulares. Sin embargo,
el agrupamiento de los miofilamentos en las miofibrillas no
es completo, y los cortes transversales muestran que las
miofibrillas están rodeadas y delimitadas solo de
manera incompleta por el retículo
sarcoplasmático y el sarcoplasma. De manera
característica, las mitocondrias son grandes,
alrededor de 2.5um de longitud y presentan crestas agrupadas
de manera compacta.Los túbulos T son invaginaciones del
sarcolema. Estos túbulos son una extensión del
espacio extracelular y poseen material de la lámina
basal extracelular que se continúa con la del
sarcolema. El retículo sarcoplásmico consta de
túbulos longitudinales e interconectados que se
acoplan con un túbulo T en la línea Z, pero no
presenta cisternas terminales grandes.Los discos intercalados son uniones celulares
especializadas situadas en las líneas Z. Si se
pudieran separar dos células en el disco, las
superficies opuestas mostrarían un diseño
complejo de papilas y crestas romas con orificios y surcos
recíprocos. Suelen cruzar una fibra a manera de
escalera por lo que tienen partes transversales y
longitudinales, estas últimas paralelas al eje de la
fibra y por lo tanto al eje de contracción. En las
regiones transversales hay demosomas en mancha o
máculas adherentes esparcidas, posiblemente para hacer
más firme la adherencia celular, y uniones intermedias
ó fascias adherentes extensas. En las regiones
longitudinales en particular hay uniones comunicantes o nexos
extensos en las regiones transversales; estos permiten una
rápida conducción del impulso entre las
células.
4.3 Contracción
El mecanismo de la contracción es de filamento
deslizante. En el adulto, en diversas regiones del corazón
las células del músculo cardiaco están
modificadas para formar el sistema de conducción de
impulsos, que regula los latidos del corazón. La
transmisión de impulsos se efectúa de una
célula muscular cardiaca a otra por vía de los
nexos.
Las células miocárdicas auriculares y
ventriculares muestran algunas diferencias. Las auriculares son
más pequeñas y con un sistema T poco desarrollado o
ausente. También contienen gránulos
electrónicamente densos relacionados con el aparato de
Golgi. Las fibras de Purkinje son células especializadas
del músculo cardiaco y forman parte del sistema de
conducción de impulsos, contienen grandes cantidades de
glucógeno; y están situadas justo por debajo del
endocardio, en la superficie interna del corazón,
particularmente en relación con el tabique
interventricular. Estas fibras forman una red compuesta de
unidades celulares independientes.
4.4 Tejido Conectivo
No es muy notable, pero se extiende entre las fibras
como un endomisio fino que contiene una red capilar muy
abundante. Los capilares linfáticos son numerosos, y a
veces se pueden ver nervios autónomos finos.
4.5 Regeneración
Después del parto, el corazón solo crece
por aumento de tamaño de cada célula muscular
cardiaca, debido a la mitosis en la vida intrauterina. En el
adulto las células alcanzan un diámetro de unos
15um, pero en condiciones patológicas (o en casos de
entrenamiento muy intensivo y prolongado) con mayor sobrecarga
cardíaca se produce hipertrofia del
corazón con aumento de la masa muscular. Esta se debe al
incremento del espesor de las fibras hasta casi 20um y
también por aumento de la longitud de cada fibra.El
corazón carece de capacidad regenerativa,
debido a la falta de capacidad mitótica en las
células musculares cardíacas después del
nacimiento y a que no existen células satélites,
como en la musculatura esquelética.
Músculo
liso
También se llama músculo no estriado o
involuntario. Su distribución es principalmente visceral,
y forma la porción contráctil de la pared del
aparato digestivo desde la parte media del esófago al ano,
incluyendo los conductos de las glándulas relacionadas con
este aparato. También se encuentran en los aparatos
respiratorios, urinario y genital; en las arterias, las venas y
los linfáticos mayores; en la dermis y en el cuerpo ciliar
del ojo. En estos lugares regula y conserva el diámetro de
la luz de las vísceras huecas.
Características del Músculo
Liso
Organización
Página siguiente  |